

Шероховатая эндоплазматическая сеть
Шероховатая ЭПС получила свое название из-за множества мелких темных, округлых гранул, расположенных на наружной, обращенной к цитозолю стороне мембраны - рибосом. Поэтому при световой микроскопии гранулярная эндоплазматическая сеть выглядит в виде базофильного вещества, дающего положительную окраску на РНК.
Шероховатая ЭПС образована уплощенными цистернами или узкими каналами, ширина полостей которых варьирует от 20 нм до нескольких микрометров (рис.11). Рибосомы на поверхности мембран ЭПС расположены в виде полисом - множество рибосом, объединенных одной информационной РНК. К мембранам они прикрепляются своей большой субъединицей.
Степень развития шероховатой ЭПС в разных клетках существенно различается. В недифференцированных клетках или клетках с низкой метаболической активностью она может быть представлена в виде редких, разрозненных мембран. Напротив, для клеток, активно синтезирующих секреторные белки, характерны локальные скопления уплощенных цистерн, называемые эргастоплазмой. Наиболее отчетливо эргастоплазма выявляется в клетках печени и поджелудочной железы, где плотно упакован ные друг около друга мембранные цистерны занимает основную часть цитоплазмы.
От функциональной активности клеток зависит также и количество рибосом, связанных с мембранами ЭПС: оно возрастает при стимуляции деятельности клетки и уменьшается при ослаблении активности внутриклеточных процессов; при дифференцировке, различных патологических состояниях клеток и т.д.
Рибосомы, связанные с мембранами ЭПС, участвуют в синтезе трех основных групп белков: секреторных белков; интегральных мембранных белков; специфических белков внутренней фазы самой ЭПС, комплекса Гольджи и лизосом.
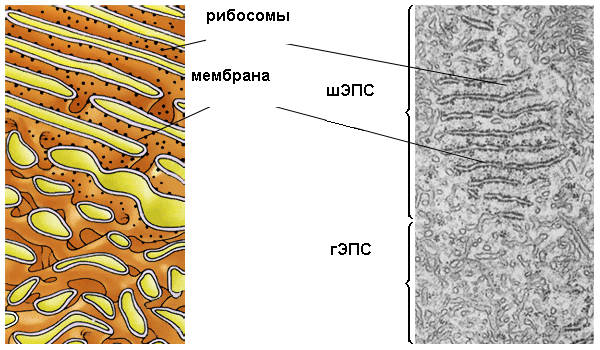
Рис. 11.Схема и электронная микрофотография шероховатой и гладкой ЭПС
Секреторные белки - это вещества, необходимые для работы других клеток или для выполнения функций, протекающих на уровне целостного организма. Такие «экспортируемые» белки выделяются, например клетками поджелудочной железы (ферменты, участвующие в расщеплении пищи в кишечном тракте); клетками печени (альбумины крови); плазмацитами (γ-глобулины) и др.
Синтез белков, предназначенных для секреции, а также молекул, необходимых для ЭПС, комплекса Гольджи, лизосом, начинается на рибосомах в цитозоле Доставка таких белков в ЭПС начинается еще до того, как полипептидная цепь полностью синтезирована, т.е. происходит одновременно с трансляцией (котрансляционно). Причем в ЭПС переносятся только те секреторные белки, которые имеют специфические сигнальные пептиды, образованные гидрофобными аминокислотами. Сигнальный пептид узнается особой сигнал-распознающей частицей, состоящей из шести полипептидных цепей и молекулы РНК. Эта частица одним своим концом соединяется с синтезированной полипептидной цепью, другой – с рибосомой, что приводит к временной остановке роста полипептидной цепи. Трансляция возобновляется после того, как сигнал-распознающая частица связывается с рецептором - интегральным белком, расположенном на цитозольной поверхности мембраны ЭПС. В результате частица отделяется и запускается АТФ-зависимый насос, который проталкивает полипептидную цепь через мембрану. Растущая цепочка вместе с сигнальной последовательностью оказываются внутри полости цистерны шероховатой ЭПС, сигнальная последовательность затем отрезается специальной протеазой, связанной с мембраной ЭПС.
Растворимые белки целиком проникают в полость ЭПС. Здесь они подвергаются посттрансляционной модификации, в ходе которой формируются дисульфидные мостики, вырезаются участки молекулы, присоединяются различные молекулы. В цистернах шероховатой ЭПС также осуществляется сворачивание вновь синтезированных белков, так, что гидрофобные остатки аминокислот оказываются в центре глобулы, а гидрофильные остатки обращены к водной среде. Для правильной укладки белка используется энергия АТФ.
Большинство растворимых белков, синтезированных в шероховатой ЭПС подвергается гликозилированию. Присоединение олигосахаридных компонентов катализируется ферментами, связанными с мембранами ЭПС. Эта реакция осуществляется путем переноса углеводного предшественника к остатку аспарагина белковой молекулы, расположенному в полости ретикулума. Разнообразие олигосахаридных структур образованных гликопротеинов возникает в результате химической модификации исходного предшественика.
Для синтеза белков цитозоля, а также органелл, не входящих в вакуолярную систему цитоплазмы, используются те же рибосомы, что и для белков, поступающих в ЭПС. Но, если синтезированный на рибосомах белок не имеет сигнального пептида для ЭПС, то образующаяся полирибосома и ее белковый продукт остаются в цитозоле. Большая часть таких белков является постоянным компонентом цитозоля, они не имеют специфических сигналов сортировки. Другие белки, содержащие сигналы сортировки, направляются в митохондрии, хлоропласты, ядра и пероксисомы, Перенос этих белков в указанные органеллы, в отличие от переноса через мембрану ЭПС, происходит после окончания трансляции.
Синтезированные и первично модифицированные в ЭПС растворимые белки транспортируются далее в аппарат Гольджи. При этом в зоне, приближенной к аппарату Гольджи, дистальные участки гранулярной ЭПС теряют рибосомы и образуют мембранные выступы, от которых отпочковываются мелкие вакуоли с белками. Отделившиеся от ЭПС вакуоли сливаются друг с другом и переносятся с помощью микротрубочек к аппарату Гольджи, где и сливаются с его мембранами.
Трансмембранные белки, предназначенные для мембраны ЭПС или других клеточных мембран, переносятся через мембрану, но не высвобождаются в полость ЭПС. Синтез мембранных белков в гранулярной ЭПС на начальных стадиях похож на синтез растворимых белков. Отличительной чертой является наличие в цепи синтезирующегося мембранного белка одной или нескольких аминокислотных стоп-последовательностей, которые препятствуют белковой цепи пересекать мембрану, в результате чего весь белок остается встроенным в мембрану. Интегральные белки всегда встраиваются в ЭПС со стороны, обращенной к цитозолю, поэтому мембрана ЭПС оказывается асимметричной.
Как и растворимые, мембранные белки могут подвергаться различным модификациям. В частности, здесь происходит первичное гликозилирование — ковалентное связывание белковой цепи со сложным олигосахаридом с помощью фермента, являющегося интегральным белком мембран ЭПС.
Важнейшая функция гранулярной ЭПС заключается в образованиии клеточных мембран. Здесь помимо синтеза на рибосомах основной части мембранных белков клетки происходит синтез большинства липидных компонентов мембран. С помощью ферментов, встроенных в мембрану, в ЭПС синтезируются фосфолипиды и холестерол. Синтез фосфолипидов мембран протекает в три этапа при участии ферментов, активные центры которых обращены в цитозоль, где находятся необходимые метаболиты. Таким образом, и образование исходных предшественников различных фосфолипидов, и их модификация осуществляются только на одной половине липидного бислоя. Для перемещения на внутреннюю сторону мембраны синтезированных во внешнем монослое липидов требуются особые белковые переносчики фосфолипидов, которые специфичны для каждого их типа. В результате бислой новообразующейся мембраны растет равномерно.
Доставка синтезированных в ЭПС фосфолипидов к мембранам органоидов, не принадлежащих к вакуолярной системе цитоплазмы, осуществляется с помощью специальных водорастворимых белков. Такие белки-переносчики захватывают молекулу фосфолипида из мембраны шероховатой ЭПС и высвобождают ее в новый липидный бислой. С помощью такого механизма фосфолипиды переносятся, например, к митохондриям и пероксисомам.
В шероховатой ЭПС синтезируется и церамидный компонент сфингофосфолипидов и сфингогликолипидов клеточных мембран. Церамид переносится в комплекс Гольджи, где к нему присоединяется фосфохолин и образованием сфингомиелина или олигосахариды с образованием гликолипидов. Ферменты, участвующие в синтезе этих соединений, расположены на нецитозольной стороне липидного бислоя.
Благодаря росту липидного бислоя увеличивается поверхность мембран вакуолей и цистерн ЭПС. Поскольку этот процесс идет одновременно с синтезом интегральных мембранных белков, мембрана, как таковая, строится и растет за счет двух процессов: синтеза и встраивания липидов, а также синтеза и интеграции мембранных белков. В дальнейшем эти мембраны становятся компонентом гладкой ЭПС, аппарата Гольджи, лизосомы или плазматической мембраны. Тип образующегося нового участка мембраны зависит от того, какие интегральные и периферические белки будут синтезироваться на рибосомах ЭПС и какие фосфолипиды будут здесь синтезироваться и далее включаться в мембрану.