

Поверхностный аппарат клеток Плазматическая мембрана
Поверхностный аппарат – неотъемлемая часть всех клеток. С его помощью осуществляются как универсальные, свойственные всем клеткам, функции (барьерная, метаболическая, транспортная и рецепторная), так и специфические функции (механическая и тургорная у клеточной стенки растений и грибов). В состав поверхностного аппарата эукариот входят плазматическая мембрана, надмембранный комплекс и субмембранная часть (кортикальный слой цитоплазмы).
Плазматической мембраной, или плазмалеммой называется часть поверхностного аппарата, окружающая каждую живую про- и эукариотическую клетку. Хотя факты о существовании плазматической мембраны появились еще в середине 19 века, ее химическая природа и строение были изучены значительно позже. Наиболее заметный вклад в изучение свойств и функций мембран внесли в 19 веке работы физиолога X.фон Моля и ботаника К.фон Негели.
Раскрытию химической природы мембран в значительной степени способствовали работы Э.Овертона, проведенные в конце 19-го – начале 20-го столетия. Им обнаружена зависимость между растворимостью различных соединений в липидах и их способностью проникать через мембраны. Данные о более высокой проницаемости мембран для жирорастворимых веществ явились первыми свидетельствами того, что они состоят из липидов, образующих непрерывный слой.
Впервые идею о двойном липидном слое высказали в 1925 году Гортер и Грендел. Эта концепция получила дальнейшее развитие в модели "сэндвича" Даниелли и Дэвсона (1935), согласно которой поверхность липидного бислоя покрыта белками. В названии модели отражается взаиморасположение слоев в мембране: липидный бислой заключен между двумя слоями белка, как масло между двумя ломтиками хлеба. Модель была подтверждена в дальнейшем экспериментами и хорошо согласовывалась с данными электронной микроскопии, изображавшими мембрану в виде своеобразных "рельсов". Вместе с тем в 60-е годы появились данные, прямо или косвенно противоречащие этой модели. Так, например, с помощью электронного микроскопа методом замораживания-скалывания внутри мембран были обнаружены глобулярные белки, целиком или частично встроенные в мембрану. При изучении мембран из различных клеточных структур показано колебание их диаметра, выявлены ферментативные свойства, найдены углеводы и т.д. Было установлено также, что некоторые белковые молекулы могут свободно диффундировать в плоскости мембраны, свидетельствуя о текучести липидного матрикса. Все эти исследования послужили базой для создания новых моделей, из которых наиболее адекватное представление о структурной организации мембран дает жидкостно-мозаичная модель, предложенная в 1972 году Синджером-Николсоном (рис. 5).
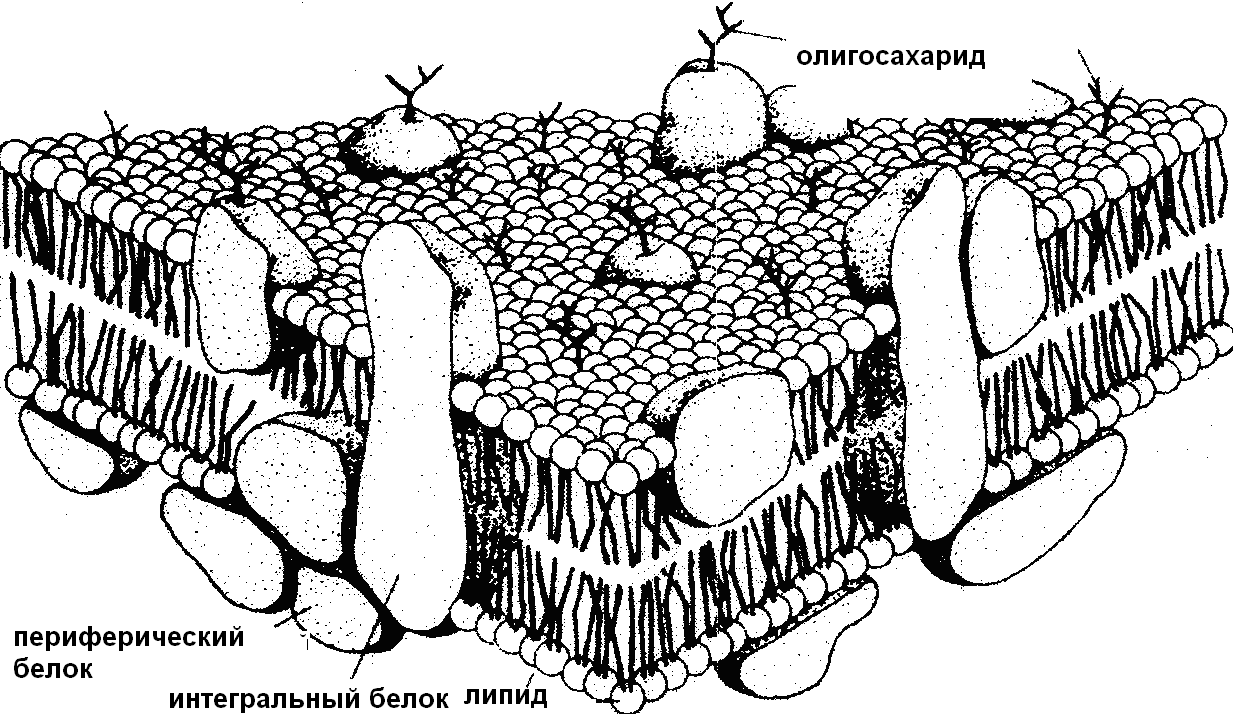
Рис. 5. Жидкостно-мозаичная модель мембраны
Согласно этой модели мембрана представляет собой текучий бислой фосфолипидов, жирнокислотные части которых направлены внутрь мембраны, а полярные головки – на ее поверхность. В него, как в мозаику, включены белки двух типов: периферические на поверхности мембраны и интегральные, погруженные в нее или прошивающие насквозь. В соответствии с функциональной специализацией мембран базовая модель бислойной структуры с погруженными в нее белками может иметь те или иные модификации.
Основными компонентами мембран являются белки и липиды. Плазматическая и некоторые другие внутриклеточные мембраны содержат, кроме того, углеводы, которые всегда являются составной частью более сложных молекул: гликопротеидов или гликолипидов. В плазматических мембранах из разных источников отношение белок/липид также может различаться, но в большинстве животных клеток весовое соотношение белков и липидов составляет 1:1.
Относительное содержание химических компонентов в других типах мембран довольно сильно варьирует и определяется в первую очередь особенностями функциональной деятельности мембран. Метаболически активные мембраны, в которых интенсивно протекают катализируемые ферментами реакции, а также мембраны, участвующие в транспортных процессах, характеризуются более высоким содержанием белка. Так, во внутренней мембране митохондрий, содержащей цепь переноса электронов, на долю белка приходится до 75% сухого вещества, тогда как в миелине, выполняющем функцию изолятора, – только 25% (табл.). Наибольшее содержание углеводов выявлено в плазматической мембране, здесь они могут составлять до 10% массы мембран.
Таблица
Соотношение белков и липидов в разных типах мембранах
Мембрана |
Соотношение белок/липид |
Миелин |
0,23 |
Плазматическая мембрана клеток печени мыши |
0,85 |
Мембрана эритроцитов человека |
1,1 |
Плазматическая мембрана амебы |
1,3 |
Плазматическая мембрана палочки сетчатки глаза |
1,0 |
Наружная митохондриальная мембрана |
1,1 |
Мембрана саркоплазматического ретикулума |
2,0 |
Внутренняя митохондриальная мембрана |
3,2 |
Липиды в плазматической мембране представлены фосфолипидами, стеролами и гликолипидами. Фосфолипиды – самые распространенные мембранные липиды, могут быть построены на основе спирта глицерина или сфингозина. В состав фосфолипидов обязательно входит молекула фосфорной кислоты (их в некоторых фосфолипидах может быть больше одной), связанная с азотсодержащим веществом или полиолом, и один или несколько жирнокислотных остатков. Различное сочетание в молекуле этих химических компонентов определяет относительно большое многообразие фосфолипидов. В эукариотических клетках чаще других фосфолипидов встречается фосфатидилхолин.
Фосфатидилхолин

Типичная молекула фосфолипидов имеет полярную головку из гидрофильных молекул и два гидрофобных углеводородных хвоста жирных кислот. Как правило, в животных клетках один из хвостов содержит насыщенную, другой – ненасыщенную жирную кислоту. Причем в фосфолипидах и других мембранных липидах преобладают жирные кислоты с ацильной цепью, составленной из 16-20 углеродных атомов. Амфипатический характер фосфолипидов, связанный с присутствием в молекуле гидрофильного и гидрофобного концов, обусловливает их способность к спонтанному формированию бислоев. Они, в свою очередь, самопроизвольно замыкаясь сами на себя, могут образовывать отдельные компартменты, что, в конечном счете, и определяет важнейшую роль фосфолипидов в формировании основного каркаса клеточных мембран.
Важнейшим свойством липидого бислоя, обусловливающим многие функции мембран, является текучесть или способность отдельных молекул свободно диффундировать в нем. Текучесть мембран зависит от многих факторов, в том числе от длины углеводородного хвоста фосфолипидов, их насыщенности, свойств и размеров гидрофильной головки.
Стеролы, состоящие из жесткого углеводородного ядра и гидроксильной группы, содержатся в мембранах клеток животных, растений и некоторых микроорганизмов. В клетках животных встречается холестерин, растений – фитостеролы, в мембранах некоторых эукариотических микроорганизмов, например дрожжей – эргостерол. Холестерин, содержащийся в плазматических мембранах животных клеток в достаточно высоких количествах, увеличивает упругость и механическую прочность липидного бислоя. Он уменьшает его текучесть и проницаемость для малых водорастворимых молекул, но в то же время, при компактном расположении жирнокислотных цепей холестерин раздвигает их, переводя бислой в более жидкую, текучую систему.
Гликолипиды – липиды, связанные с углеводами, построены, как и фосфолипиды, на основе спирта глицерина (у растений и бактерий) и сфингозина (у животных). У гликосфинголипидов углеводы присоединены с помощью гликозидной связи к концевой гидроксильной группе церамида, состоящего из молекулы сфингозина и жирнокислотного радикала. Церамид гликосфинголипидов, обладающий гидрофобными свойствами, погружен в липидную фазу мембран, а углеводный компонент ориентирован наружу в окружающую полярную среду.
Церамид

В плазматической мембране животных клеток, где на их долю приходится от 2 до 10% массы всех липидов, гликолипиды могут содержать как остатки моно-, так и олигосахаридов, и иметь достаточно сложное строение. Важная роль гликолипидов сводится к их участию в процессах межклеточного взаимодействия, принятию сигналов. Так, в нервной системе они участвуют в синаптической передаче и играют особую роль при формировании межнейрональных связей.
Белки мембран подразделяют на периферические, расположенные на поверхности липидного бислоя, и интегральные белки, частично или полностью пронизывающие бислой. Периферические белки удерживаются поверхностью мембраны преимущественно за счет слабых электростатических взаимодействий с другими белковыми молекулами и с полярными головками липидов. Интегральные белки погружены в толщу липидного бислоя или пронизывают мембрану насквозь. Такие белки, как и окружающие их липиды, амфипатичны, их трансмембранные участки (домены), образующие внутри мембраны -спираль из гидрофобных аминокислот, связаны с неполярной областью липидного бислоя, а гидрофильные участки обращены с обеих сторон мембраны к воде. Большая часть трансмембранных белков на внеклеточной стороне плазматической мембраны связана с олигосахаридами, т.е. относится к гликопротеинам. Мембранные белки чрезвычайно разнообразны и, в конечном счете, именно они определяют функциональную активность и уникальность плазматических мембран из различных источников.
К числу периферических белков относится мембранный скелет клеток, расположенный на обращенной к цитоплазме стороне плазматической мембраны. Во многих эукариотических клетках цитоскелет обеспечивает механическую опору для плазматической мембраны, определяет форму клетки. Некоторые периферические белки, как на внутренней, так и на внешней стороне мембраны обладают ферментативной активностью. В частности, с внутренним монослоем плазматической мембраны связаны ферменты гликолитического цикла, а с внешним – катаболические ферменты, в том числе ацетилхолинэстераза, осуществляющая гидролиз ацетилхолина.
Интегральные белки могут выполнять функции каналов или переносчиков для ионов и некоторых молекул, выступать в качестве ферментов, обеспечивающих энергозависимый трансмембранный перенос ионов. Некоторые интегральные белки являются частью сложного рецепторного комплекса для приема гормональных сигналов, выполняют функции узнающих элементов в мембранном транспорте.
Углеводы в свободном состоянии в мембранах практически не встречаются. Они ковалентно присоединены к мембранным белкам (гликопротеины) и к липидам (гликолипиды). В составе сложных молекул углеводы могут быть представлены одной молекулой (как, например, в цереброзидах) или же – олигосахаридной цепочкой. Помимо нейтральных моноз, таких, как глюкоза, галактоза и фукоза углеводные компоненты гликолипидов и гликопротеинов содержат производные моносахаридов, обладающие основными (гексозамины) или кислыми (сиаловые кислоты) свойствами. Углеводы обеспечивают процессы, протекающие на поверхности плазматической мембраны, они определяют отрицательный заряд поверхности клеток, препятствующий агрегации свободно циркулирующих клеток крови.
Надмембранные структуры поверхностного аппарата подразделяются на собственно надмембранный комплекс, или гликокаликс и его производные (клеточные стенки). В клетках разных типов они выполняют общие для всех клеток функций, но могут нести и особые функциональные нагрузки, например, механическую, тургорную, локомоторную и др. В связи с этим компоненты надмембранного комплекса весьма многообразны по своей структурной организации и химическому составу.
Под гликокаликсом подразумевается обогащенная углеводами периферическая зона поверхности большинства эукариотических клеток (рис. 6). Основная часть углеводов гликокаликса присоединена к молекулам белков и липидов плазматической мембраны. Им наряду с белками принадлежит важная роль в межклеточном узнавании и узнавании между клеткой и матриксом. Наиболее развитый гликокаликс встречается в нервных клетках: здесь углеводы, входящие в состав сложных гликолипидов – ганглиозидов, обеспечивают в синаптических мембранах рецепцию медиаторов, участвуют в формировании межнейрональных связей и т.д. С углеводами связано наличие специфических антигенов на поверхности эритроцитарных мембран.

Рис. 6. Электронная микрофотография поверхности клетки с гликокаликсом
1 – гликокаликс; 2 – плазматическая мембрана; 3 – цитоплазма; 4 – ядерная оболочка; 5 – хроматин
В примембранной зоне помимо углеводов находятся белки, выполняющие многообразные функции. Так, внешние участки интегральных белков участвуют в рецепции различных сигнальных молекул и обеспечивают клеточную адгезию. Они формируют рецепторы главного комплекса гистосовместимости клеток разных тканей, определяющих уникальность и неповторимость поверхности клеток не только у разных видов животных, но и у каждого индивидуума.
Кроме компонентов, непосредственно входящих в состав плазматической мембраны, гликокаликс может включать в себя вещества, которые секретируются самими клетками и затем адсорбируются на клеточной поверхности. Здесь локализуются, в частности, гидролитические ферменты, которые выделяются клеткой и связываются с зоной гликокаликса с помощью ионов кальция и магния.
С надмембранным комплексом, тесно контактируют такие углеводсодержащие макромолекулы внеклеточного матрикса, как гликопротеины и протеогликаны - сложные высокомолекулярные комплексы белков и полисахаридов гликозаминогликанов. Учитывая этот факт, следует понимать, что не существует четкой границы между компонентами плазматической мембраны и адсорбированными на ней макромолекулами внеклеточного матрикса.
И, наконец, в состав гликокаликса могут входить соединения, которые самой клеткой не производятся, а лишь адсорбируются ее поверхностью. Известно, что с гликокаликсом микроворсинок кишечного эпителия связываются ферменты из кишечной полости, участвующие в процессах пристеночного пищеварения. Особый тип надмембранных структур эукариотических клеток - это клеточные стенки растений и грибов. Основными компонентами этих производных надмембранного комплекса являются линейные полисахариды, представленные у растений целлюлозой (полимером глюкозы), а у многих групп грибов – хитином (полимер N-ацетилглюкозамина). Клеточные стенки выполняют функцию наружного механического каркаса (скелета) клеток, защищают ее поверхность, обеспечивают тургорные свойства.
Клеточные стенки принципиально сходны по своему строению: состоят из опорного каркаса из параллельно расположенных волокон целлюлозы или хитина, связанных поперечными перемычками. Промежутки между волокнами заняты аморфным гелеобразным матриксом с большим содержанием воды. В состав матрикса у растений входят полисарахиды гемицеллюлозы и пектиновые вещества. Соотношение между количественным содержанием волокнистых и аморфных компонентов стенки может сильно варьировать у различных представителей растительного мира и в разных органах растений. Часто для придания клеточным стенкам особых свойств (в частности, для повышения прочности) аморфный матрикс дополнительно пропитывается органическими веществами, как например лигнином, или укрепляется минеральными веществами и др.
У грибов матрикс клеточной стенки представлен не углеводами, а белками, в том числе ферментами, участвующими во внеклеточном расщеплении биополимеров.
Фибриллы целлюлозы собираются из моносахаридов не внутри клетки, а на ее поверхности с помощью специальных ферментов, встроенных в плазматическую мембрану. Причем ориентация волокон целлюлозы соответствует расположению белковых элементов опорно-двигательного аппарата клетки, расположенного непосредственно под плазматической мембраной. Аморфные вещества матрикса, гемицеллюлозы и пектиновые вещества синтезируются внутри клетки и выделяются из нее через вакуоли аппарата Гольджи путем экзоцитоза. Таким образом, клеточные стенки можно рассматривать как продукт жизнедеятельности клетки.
Субмембранная, т.е. обращенная к цитоплазме часть поверхностного аппарата эукариотических клеток, образована тесно примыкающей к плазматической мембране сетью фибриллярных белков. Эти ассоциированные с липидным бислоем белки рассматриваются одновременно и как скелет плазматической мембраны и как кортикальный слой цитоплазмы, т.е. элемент цитоскелета. В этом смысле разграничение понятий скелет мембраны и цитоплазмы, как и в случае составных компонентов гликокаликса, является достаточно условным. В организации скелета участвует целый ряд периферических белков. Наиболее полно структура и функции субмембранных компонентов плазматической мембраны изучены на примере эритроцитов (рис. 7). Мембраны эритроцитов или «тени» легко получить, если поместить клетки в гипотонический солевой раствор, вызывающий их набухание и лизис. Если далее растворить мембранные тени в определенных условиях, то получается препарат белков, составляющих скелет мембраны эритроцитов.
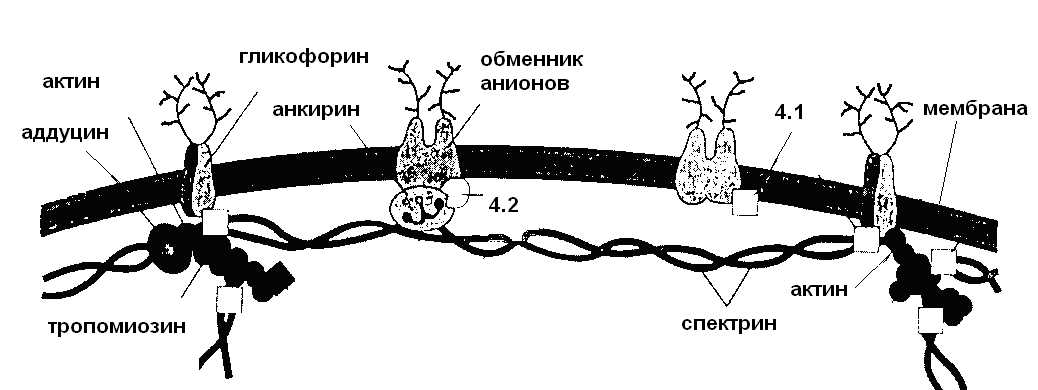
Рис. 7. Субмембранный комплекс эритроцитов
Самый распространенный белок, поддерживающий структурную целостность и дисковидную форму эритроцитов – это спектрин. Спектрин представляет собой тетрамер, в котором мономеры, связанные в положении «голова к голове», формируют тонкий филамент. Окончания спектриновых филаментов соединяются с актином - важнейшим белком опорно-двигательного аппарата большинства эукариотических клеток, а также с рядом других белков. Прикрепление этого комплекса к интегральным белкам мембраны обеспечивается различными связывающими белками. Подобные или сходные по строению и функциям белки, образующие субмембранную трехмерную сеть филаментов, найдены во многих типах клеток, где они определяют форму и механические свойства плазматической мембраны.
В субмембранном комплексе сосредоточены ферментные системы, связанные с процессами рецепции. В частности, здесь осуществляется образование некоторых вторичных посредников, через которые происходит передача внутрь клетки сигналов поступающих в нее со стороны надмембранного аппарата. Таким образом, занимая пограничное положение между плазматической мембраной и метаболическим аппаратом клетки, субмембранный комплекс отвечает за реализацию клеточных ответов на гормональные и другие сигналы, реакцию клетки на изменения в окружающей среде.