

Микротрубочки
Микротрубочки обнаружены во всех эукариотических клетках. Они представляют собой длинные полые цилиндры с внешним диаметром около 25 нм и длиной, достигающей несколько мкм. Как уже отмечалось выше, функции микротрубочек довольно многообразны: формирование поддерживающего каркаса клетки; участие в расхождении хромосом при делении клетки и в упорядоченном внутриклеточном транспорте гранул, везикул и органелл; обеспечение подвижности клеток (из микротрубочек образованы реснички, жгутики, базальные тельца и центриоли).
Микротрубочки построены из основного структурного белка тубулина и ассоциированных с ним белков. При полимеризации тубулина образуются тонкие нити – протофиламенты, которые скручиваются в полую трубку (рис. 29).
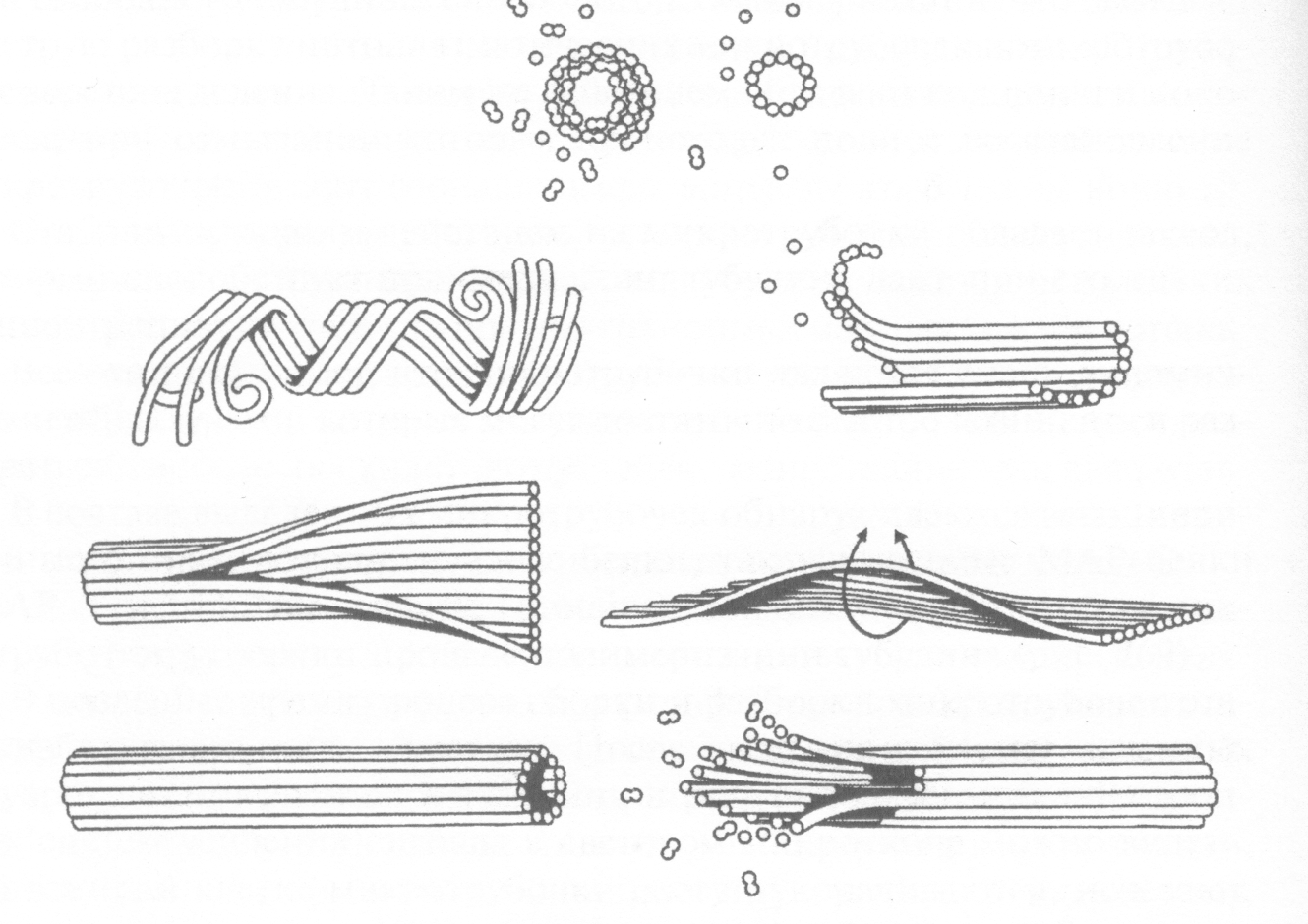
Рис. 29. Схема самосборки микротрубочек
Каждая молекула тубулина является гетеродимером, состоящем из α и β субъединиц. По окружности микротрубочки уложено 13 (или 11, как, например в В-трубочке ресниченк и жгутиков) таких тубулиновых протофиламентов. В отличие от микрофиламентов полимеризация тубулина идет в присутствие не АТФ, а ГТФ, причем с ГТФ связаны обе субъединицы, а гидролизу подвергается только молекула на β субъединице.
Микротрубочки полярны, при достаточной концентрации тубулина в окружающей среде димеры, связанные с ГТФ, быстро добавляются к растущему плюс-концу (рис. 30). На минус-конце происходит отщепление димеров. Сборку микротрубочек могут останавливать вещества, которые влияют на полимеризацию тубулина. Так, алкалоид колхицин из безвременника осеннего прочно связывается со свободными молекулами тубулина и препятствует тем самым их включению в растущую микротрубочку. Например, в результате снижения концентрации свободного тубулина в цитоплазме клетки при ее делении происходит разборка микротрубочек веретена деления и останавливается расхождение к полюсам хромосом.
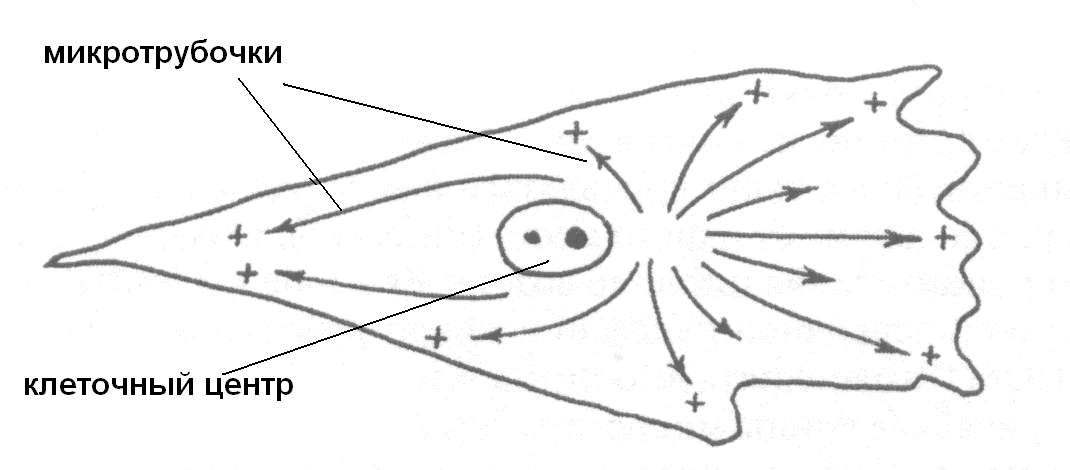
Рис. 30. Полярное расположение микротрубочек в цитоплазме
Микротрубочки очень динамичные структуры. В живой клетке происходит непрерывный процесс сборки новых микротрубочек. Начальный этап этого процесса – нуклеация происходит в особых участках клетки, называемых центрами организации микротрубочек (ЦОМТ). Здесь из тубулиновых субъединиц образуются небольшие структуры, обращенные своим минус концом к ЦОМТ. В этой зоне деполимеризация тубулина ограничена или заблокирокана с помощью специальных белков. При высокой концентрации свободного тубулина в среде происходит их полимеризация: димеры тубулина присоединяются к плюс-концу микротрубочек, который может расти очень быстро по направлению к периферической части клетки. Таким образом, ЦОМТ постоянно производит микротрубочки, растущие к периферии клетки, тогда как другие микротрубочки укорачиваются по направлению к нему. Микротрубочки, теряющие связь с ЦОМТ, могут быстро разбираться и, наоборот, стабилизируются, если присоединяются к нему своим минус-концом.
В животных клетках центры организации микротрубочек представляют собой уплотненные участки цитоплазмы, расположенные главным образом в области клеточных центров (центросом), содержащих парные цилиндрические структуры центриоли (рис. 31). Если клетку обработать веществом, деполимеризующим тубулин, то микротрубочки исчезают. При удалении этого вещества микротрубочки появляются вновь, причем растут они синхронно в направлении от клеточного центра с такой же скоростью, с которой тубулин полимеризуется in vitro – около 1 мкм/мин при 37оС. Через 75 мин радиальное расположение микротрубочек в клетке восстанавливается.

Рис. 31. Схема расположения центросом в клетках различных типов
На полярный рост микротрубочек оказывают влияние ассоциированные с ними белки, называемые МАР-белками (в переводе с английского – microtubule associated proteins). Благодаря участию этих белков в дифференцированных клетках уменьшается деполимеризация тубулинов, в силу чего микротрубочки остаются относительно стабильными достаточно длительное время (до нескольких часов). С другой стороны, стабилизация микротрубочек достигается за счет их модификации (например, ацилирования аминокислот). Во время деления клетки скорость образования и деполимеризации микротрубочек значительно возрастает, что обеспечивает соединение хромосом с микротрубочками и формирование митотического веретена.
В цитоплазме существует равновесие между полимеризованным и неполимеризованным тубулином, а в ресничках и жгутиках он встречается только в полимеризованной форме. Длина микротрубочек, их количество и распределение различается на различных стадиях клеточного цикла, во время роста аксонов и т.д.
Как и актиновые микрофиламенты, тубулины связаны с рядом вспомогательных белков, влияющих на организацию и функции микротрубочек. Некоторые из них были уже рассмотрены выше - это факторы инициации, необходимые для образования и удлинения микротрубочек. Известны ассоциированные с микротрубочками белки, которые могут образовывать своеобразные боковые шипы и выступы длиной до нескольких десятков нм. Считается, что функции этих белков заключаются в связывании микротрубочек между собой и с другими компонентами цитоплазмы или создании защитной зоны вокруг самой микротрубочки. Другой тип белков отвечает за обеспечение процессов внутриклеточного движения органелл и везикул. В ресничках и жгутиках обнаружены белки, ответственные за сцепление микротрубочек между собой и образующие сложные системы и структуры.
Важнейшей функцией микротрубочек является обеспечение ориентированного движения внутриклеточных структур. Их часто рассматривают как «рельсы», направляющие перемещения по клетке органелл. С помощью такого механизма осуществляется как перемещение цитоплазматических органелл, так и транспорт везикул вдоль аксонов нервных клеток. Направленное движение органелл останавливается при деполимеризации тубулина, что в целом нарушает жизнедеятельность клеток. В частности, составные компоненты комплекса Гольджи, обычно локализованные вблизи клеточного центра, смешиваются и оказываются разбросанными по всей цитоплазме. Промежуточные филаменты при этом перемещаются во внутреннюю часть клетки и образуют клубок вокруг ядра.
Внутриклеточное движение опосредовано особыми белками (моторные белки) и требует затрат энергии АТФ. В настоящее время известно несколько двигательных молекул, обеспечивающих перемещение митохондрий, лизосом, везикул по микротрубочкам. При этом двигательные молекулы перемещаются по микротрубочкам лишь в одном направлении. Кинезины – это белки, движущиеся от минус – к плюс концу, т.е. в направлении от ЦОМТ к периферии клетки; динеины – белки, перемещающиеся в обратном направлении: от периферии к центру клетки.
Моторные белки обоих классов имеют принципиально сходное строение и напоминают по структуре и химической природе миозины. Они состоят из двух тяжелых и нескольких легких цепей. У кинезина тяжелые цепи формируют с одной стороны хвостовой стержневой домен, который непосредственно взаимодействует с транспортируемыми внутриклеточными структурами. В связывании моторного белка с мембраной пузырьков и органелл участвуют легкие цепи кинезина (рис. 32).
С противоположной стороны тяжелых цепей расположено два головных домена, имеющих глобулярную конформацию. Каждая головка обладает АТФазной активностью и контактирует с глобулами тубулина, вдоль которого перемещаются молекулярный двигатель. При гидролизе АТФ изменяется конформация молекулы кинезина и положение головок по отношению к тубулину, в результате чего происходит перемещение кинезина вдоль микротрубочки. При движении каждая из головок кинезина связывается с микротрубочкой попеременно,т.е молекула «шагает». Длина такого шага точно соответствует размеру димера тубулина. За 1сек. кинезин совершает около 100 «шагов» и проходит примерно 800 нм.
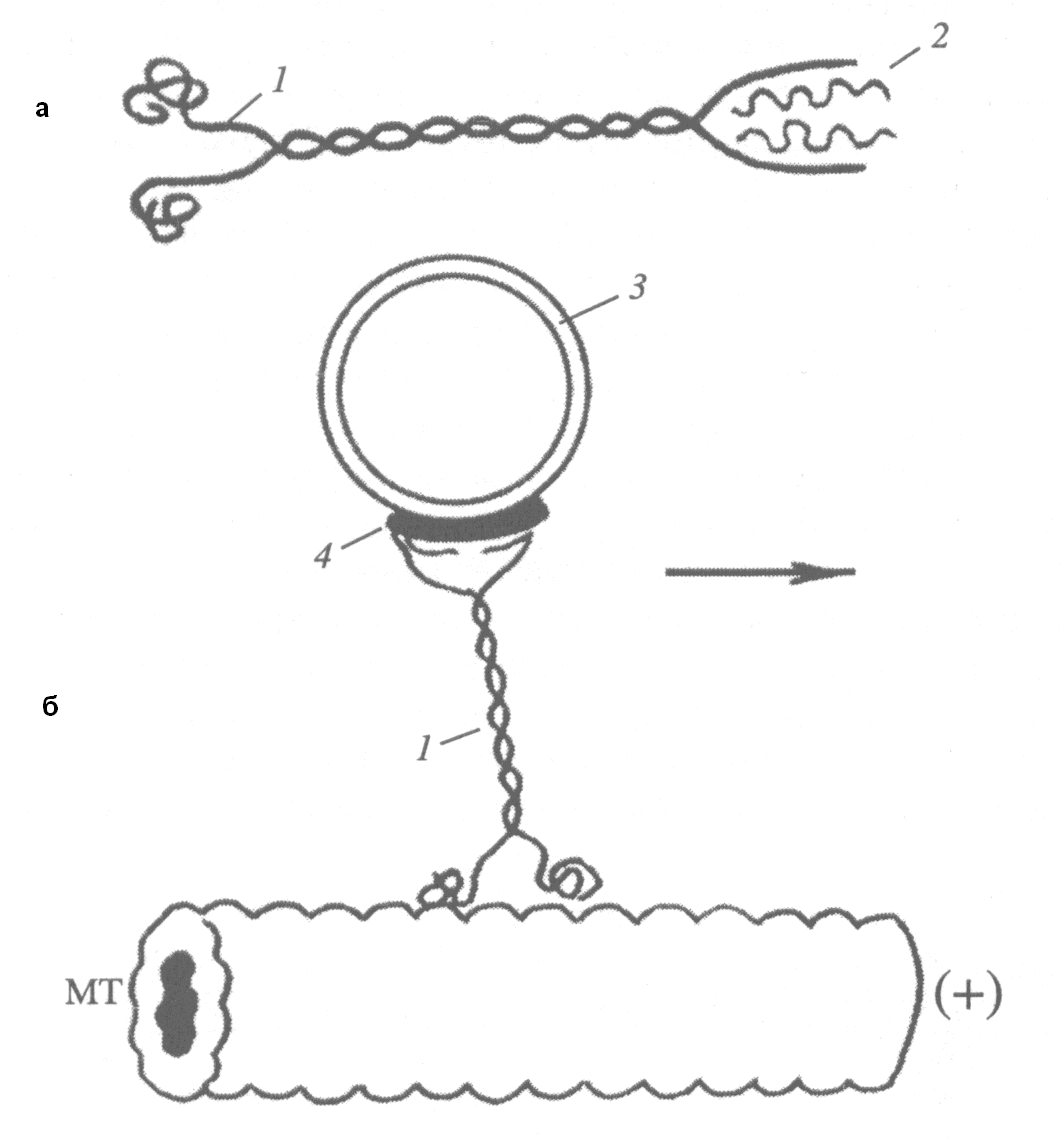
Рис. 32. Схема внутриклеточного транспорта органелл с помощью кинезина
а -строение молекулы кинезина: 1 – тяжелые цепи, 2 – легкие цепи
б – участие кинезина в транспорте: МТ –микротрубочка, 1 – кинезин, 3 – везикула, 4 - кинектин
В настоящее время описано несколько разных форм кинезинов, различающихся по строению хвостовых доменов. Некоторые из них проявляют специфичность по отношению к транспортируемым структурам, т.е. осуществляют перенос только митохондрий, синаптических пузырьков или хромосом.
Динеины имеют аналогичное строение. Головки двух его тяжелых цепей связываются с микротруточками, а промежуточные и легкие цепи – с вакуолями. Различают цитоплазматические динеины, переносящие вакуоли и хромосомы к минус концу микротрубочек, и аксонемные, входящие с состав ресничек и жгутиков и обеспечивающих их движение.
Таким образом, радиально расходящиеся от клеточного центра микротрубочки не только формируют внутриклеточный скелет, но и, благодаря строго ориентированной деятельности различных моторных белков, образуют векторную систему движения различных структур. При этом лизосомы, везикулы, отпочковывающиеся от ЭПС, транспортные и секреторные вакуоли комплекса Гольджи перемещаются от центра клетки к ее периферии, и, наоборот, эндосомы, а также везикулы ЭПС и комплекса Гольжди, подвергающиеся рециклизации, - от периферии к центру.
Микротрубочки обнаружены в эукариотических клетках не только в свободном состоянии, но и в ресничках и жгутиках подвижных клеток, где они расположены строго определенным образом: по две микротрубочки лежат в центре - центральная пара, а вокруг них располагаются девять пар других микротрубочек – периферические дублеты (рис. 33.).

Рис. 33. Схема поперечного среза реснички.
Вся эта структура, называемая аксонемой, находится под плазматической мембраной, которая является продолжением плазматической мембраны. Различают А и В микротрубочки и белки, ассоциированные с тубулином: динеин, образующий наружную и внутреннюю ручки, белки радиальной спицы и нексин. Динеиновые ручки отходят через равные интервалы от дублета микротрубочек, при этом основание молекул динеина связано с А-трубочкой дублета. На глобулярных головках динеиновых ручек находятся участки для АТФ–зависимого присоединения их к В-микротрубочке другого дублета (рис. 34).

Рис.34. Схема строения фрагмента микротрубочки реснички со связанной с ней молекулой динеина
Последовательность процессов скольжения дублетов микротрубочек изображена на рис. 35.
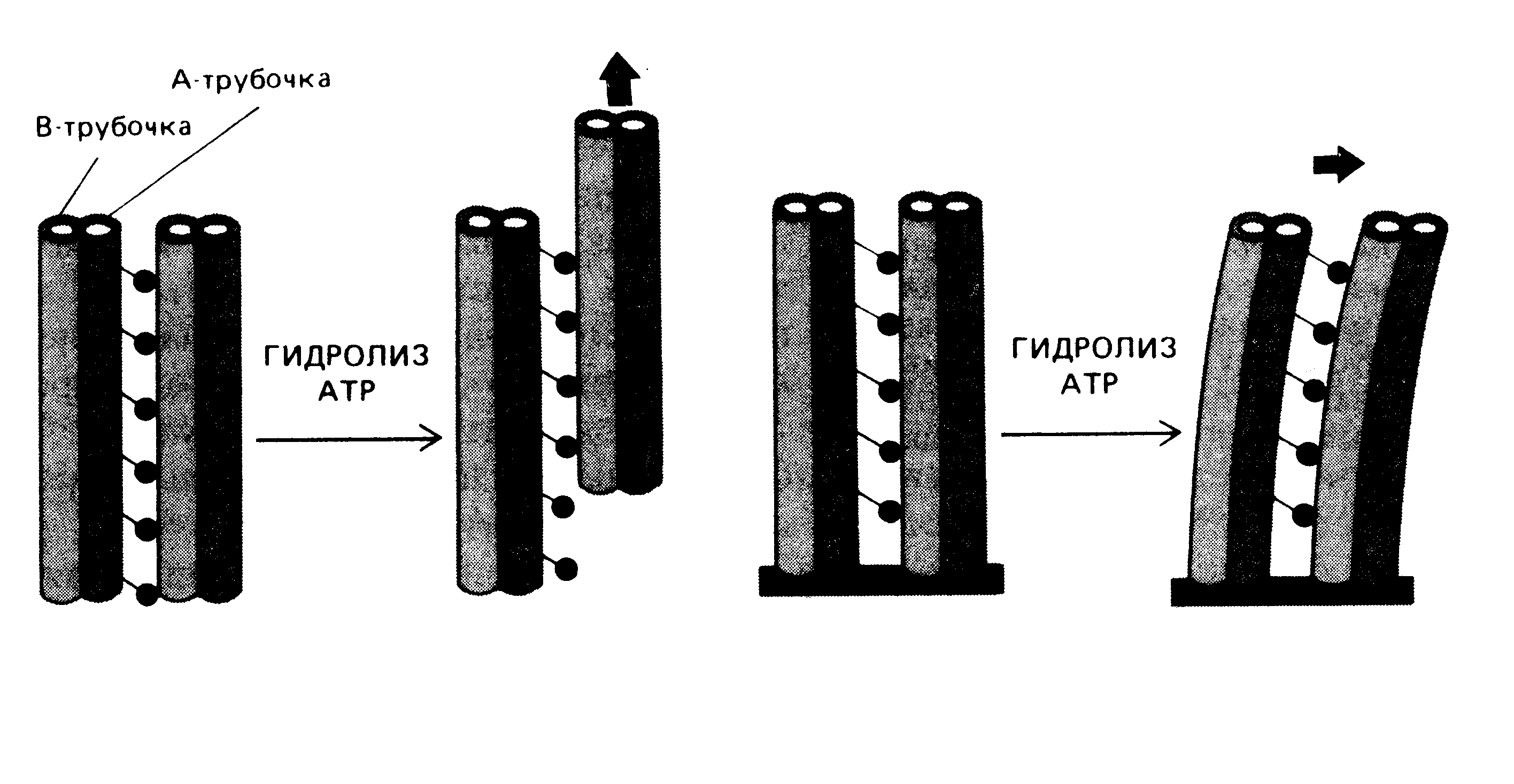
Рис. 35. Схема скольжения периферийных дублетов микротрубочек в ресничке
На первом этапе осуществляется гидролиз АТФ, что обеспечивает движение динеиновых ручек по направлению к минус-концу микротрубочки второго дублета. Далее происходит продольный сдвиг двух соседних дублетов; поперечное сшивание дублетов микротрубочек; новое скольжение дублетов, приводящее к изгибу дублетов.
Биение ресничек и жгутиков обеспечивает силы, необходимые для движения свободноживущих клеток (у инфузорий и некоторых жгутиконосцев) или для перемещения различных веществ вблизи их поверхности (например, у эпителиальных клеток дыхательных путей многих беспозвоночных и позвоночных животных). Способ движения с помощью ресничек и жгутиков может быть разнообразным: маятникообразным, воронкообразным или волнообразным.
У основания ресничек и жгутиков находятся базальные тельца – структуры, которые состоят из микротрубочек, объединенных в 9 триплетов. Базальные тельца участвуют в образовании ресничек и жгутиков.
Сходное строение имеют центриоли, являющиеся во всех животных клетках срединным элементом центросомы, или клеточного центра, который был обнаружен в конце 19 века. Как уже указывалось выше, в интерфазных клетках от клеточного центра происходит образование и рост микротрубочек. Центросомы отсутствуют у высших растений, низших грибов и некоторых простейших, в то же время они имеются у низших растений, например в зооспорах водорослей.
Обычно центросомы представлены парой центриолей (диплосома), которые расположены перпендикулярно по отношению друг к другу. Они окружены зоной более светлой цитоплазмы и радиально расходящихся фибрилл (центросфера). Центриоли способны репродуцироваться: новообразующаяся центриоль формируется рядом со старой, а затем «материнская» и «дочерняя» центриоли расходятся.
Центриоль – это небольшая цилиндрическая органелла, построенная из микротрубочек. Они образуют 9 триплетов микротрубочек, входящих в состав стенки центриолей (рис. 36). Каждый триплет содержит одну полную микротрубочку, содержащую 13 гетеродимеров тубулина (А), и две неполные – по 11 молекул тубулина (В и С). Поперечные сшивки из белковых молекул участвуют в поддержании цилиндрической структуры. От микротрубочки А отходят «ручки», одна из которых направлена к микротрубочке С соседнего триплета, а другая – к центру цилиндра.
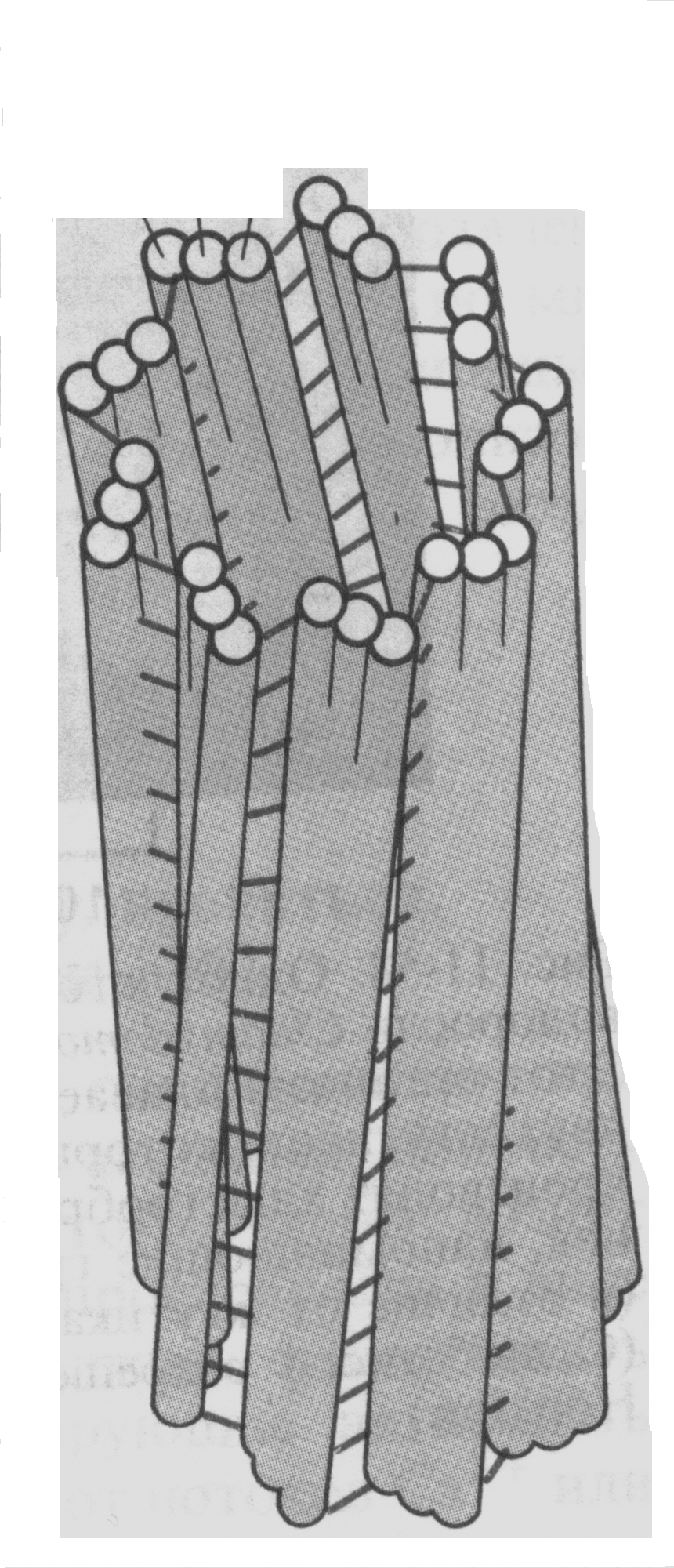
Рис. 36. Схематическое изображение центриоли