

8.1.3. Механизм расплетения спирали днк
В рассмотренной картине расплетения спирали возникает следующая проблема: при движении глазка (репликационной вилки) основные не расплетенные части цепи должны быстро вращаться. Для этого требуются, с одной стороны, значительные затраты энергии, а, с другой стороны, вращение топологически не всегда возможно. В действительности вращения всей цепи не происходит, так как специальные ферменты ДНК-топоизомеразы создают в исходной двойной цепи одно- или двухцепочечные разрывы, позволяющие цепям ДНК разделиться, не затрагивая основную цепь. При разрыве одной цепи появляется вращательная степень свободы вокруг одной из одинарных ковалентных связей другой цепи. Таким образом, движение репликационной вилки сопровождается разрывами двойных цепей под действием ДНК-топоизомеразы, которая затем, в процессе синтеза дочерней цепи, восстанавливает целостность спиралей. Разрывы цепей ликвидируются.
8.1.4. Репликоны
Эукариотические клетки перед каждым делением должны синтезировать копии всех своих хромосом, так как при делении клеток реплицируется весь геном. Поскольку время ограничено, то в процессе репликации все хромосомы разделяются на множество отдельных участков – репликонов. Репликоны активируются не одновременно, но перед каждым клеточным делением все они должны быть реплицированы. Процесс расщепления при репликации происходит одновременно во многих точках начала репликации в каждой молекуле ДНК. В каждый момент времени в хромосомах двигается, независимо друг от друга, множество репликационных вилок. Остановка вилок происходит только при достижении конца хромосомы, или при столкновении вилок, движущихся в противоположных направлениях. В результате время репликации резко сокращается.
Процесс репликации – кооперативное явление, в котором все задействованные белки взаимодействуют между собой так, что репликация является результатом их согласованного действия.
§8.2. Транскрипция
Структура полипептидной цепи белка кодируется определенными генами. Информация, записанная в гене, передается по цепи переноса информации: ДНК–РНК–белок. Процесс синтеза белков, контролируемый ферментами, структура которых также записана в генах ДНК, осуществляется в два этапа.
1. Информация, записанная в виде последовательности нуклеотидов на одной из двух комплементарных цепей в ДНК, переписывается в одно-цепочечную молекулу РНК в ядре клетки путем синтеза на ДНК комплементарной цепочки РНК. Синтезированная РНК называется информационной РНК (иРНК) или матричной (мРНК). Этот процесс называется транскрипцией (от лат. transcritio – переписывание). Транскрипция – это биосинтез мРНК на матрице ДНК. Транскрипция, так же как репликация ДНК, происходит в ядре клеток.
2. По мРНК в цитоплазме клетки строится полипептидная цепь первичной структуры белка с заданной последовательностью аминокислотных остатков. Этот процесс принято называть трансляцией (от лат. translatio – передача).
8.2.1. Инициация транскрипции
Инициация начала белкового синтеза, предшествующего делению клеток, производится специализированными белками – факторами роста, вырабатываемыми обычно другими клетками. Факторы роста с наружной стороны клетки связываются со специальными рецепторами (от лат. recipere – получать, receptor – принимающий), пронизывающими мембрану (рис. 8–3). Это приводит к активации примембранных ферментов – протеинкиназ, связанных с молекулой рецептора на внутренней поверхности мембраны. Протеинкиназа запускает каскадную реакцию фосфорилирования ряда белков: А, Б и т.д. (рис. 8–3).
При этом фосфорилирование белка А превращает его в протеинкиназу, которая фосфорилирует белок Б и превращает его в активную протеинкиназу, которая в свою очередь, специфично фосфорилирует ряд других белков. Цепь передачи сигналов заканчивается в ядре, где происходит активация (фосфорилирование) факторов транскрипции, образующих семейство J, состоящее из белков J1, J2, J3 и т.д.
Эти белки, как факторы транскрипции, активны только находясь в парном состоянии J1–J1, J2–J2, а также J1–J2, или J2–J3.
При образовании
пар во много раз увеличивается число
возможных активных димеров. Если
семейство J, состоит из n
белков, то число различных димеров равно
![]() .
.
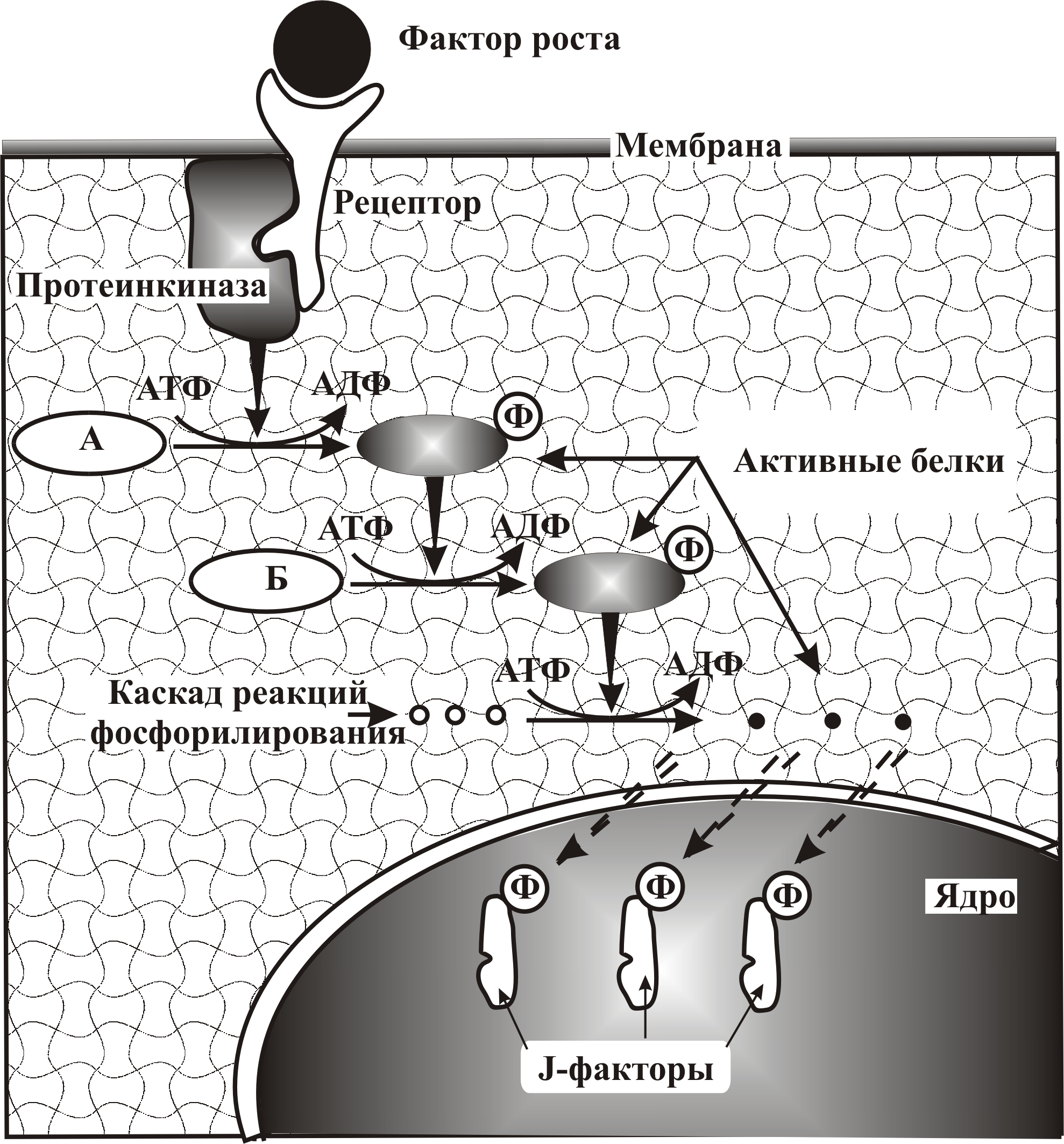
Рис. 8–3. Активация транскрипции генов. К белковой молекуле – рецептору, пронизывающему мембрану, присоединяется белковый фактор роста, управляющий делением клетки. В результате примембранный фермент – протеинкиназа, взаимодействующая с рецептором, активируется и запускает каскад реакций фосфорилирования: фосфорилиривание белка А превращает его в активную протеинкиназу, которая фосфорилирует белок Б, фосфорилирующий в свою очередь ряд других белков. Каскад фосфорилирования заканчивается фосфорилированием J-факторов транскрипции, запускающих работу генов, необходимых для клеточных делений
Заметим, что механизм запуска фосфорилирования постепенно затухает, если нет постоянных внешних сигналов, стимулирующих синтез белков необходимых для деления клеток и роста тканей. Кроме того, активность фосфорилированных димерных факторов транскрипции может быть блокирована стероидными гормонами (органическими полициклическими соединениями), легко проникающими через липидную мембрану в ядро клетки в составе гормон-рецепторных комплексов. Одним из возможных механизмов блокирования транскрипции является диссоциация (отрыв) J–J димеров, связанных с промоторами (от лат. promoveo – продвигаю) ДНК. В результате транскрипция генов подавляется.