

6.1.3. Наследственные болезни
Доминантные и. сцепленные с полом рецессивные заболевания. С точки зрения популяционной генетики доминантные и сцепленные с полом рецессивные болезни удобно разделить на две категории.
1. Болезни, при которых способность к размножению нарушена либо потому, что пораженные погибают в раннем возрасте, либо потому, что их дефекты настолько серьезны, что препятствуют вступлению в брак.
2. Болезни, при которых способность к размножению не нарушена, либо потому, что дефекты незначительны, либо потому, что проявляются они только после завершения репродуктивного периода.
Частота встречаемости и распространение заболеваний первой группы в основном определяются частотой возникновения соответствующих мутаций, т. е. скоростью мутирования: большинство мутаций такого рода исчезает из популяции через одно или несколько поколений. Этот вопрос обсуждался в разд. 5.1; данные по скорости мутирования для таких заболеваний свидетельствуют о сходстве этого показателя во всех изученных популяциях. Факторы отбора специфичны для каждой болезни и более или менее идентичны во всех популяциях, поэтому частота встречаемости заболеваний в них также сходна. Если в разных популяциях применяют методы лечения разной эффективности, частота заболевания в них также будет разной. Например, успешное лечение гемофилии приведет к увеличению числа гемофиликов в популяции.
Совершенно иная ситуация имеет место в случае болезней, не затрагивающих размножение. Частота их встречаемости в разных популяциях может быть совершенно различной в зависимости от размера популяции, ее истории и брачной структуры. Эти вопросы рассматриваются в разд. 6.4.2.
Аутосомные рецессивные заболевания. Различные факторы, определяющие частоты аутосомных рецессивных заболеваний в популяциях человека, в основном неизвестны. Поэтому невозможно точно определить, каким образом эти частоты будут меняться под влиянием современной цивилизации. Частота встречаемости большинства аутосомных рецессивных заболеваний неизвестна. При осуществлении обширных программ скрининга новорожденных недавно были получены оценки частоты нескольких врожденных нарушений метаболизма [1881].
Организаторы исследования, целью которого было определение частоты фенилкетонурии, выбирали для участия в нем те центры скринирования, которые применяют удовлетворительные методики и анализируют достаточное количество новорожденных. Данные учитывали только в том случае, если выборка обследованных составляла не менее 70000 новорожденных. Центры, обнаружившие региональные различия в пределах исследуемой популяции, должны были дать отдельные оценки для каждого района
292 6. Популяционная генетика
Таблица 6.4. Частота фенилкетонурии и гиперфенилаланинемии в некоторых популяциях [1881] |
||
Область |
Частота |
|
|
ФКУ |
Гиперфенилаланинемия |
Варшава, Польша |
1: 7782 |
1: 16885 |
Прага, ЧССР |
1: 6618 |
1: 6303 |
ГДР |
1: 9329 |
1: 52135 |
Восточная Австрия |
1: 8659 |
1: 21982 |
Западная Австрия |
1: 18 809 |
1: 18 809 |
Швейцария |
1: 16644 |
1: 24106 |
Эвиан, Франция |
1: 13715 |
1: 13 143 |
Гамбург |
1: 9081 |
1: 61297 |
Мюнстер, ФРГ |
1: 10934 |
1: 7997 |
Гейдельберг |
1 : 6178 |
1: 14 580 |
Дания |
1: 11897 |
1 : 40 790 |
Стокгольм, Швеция |
1: 43 226 |
1: 22140 |
Финляндия |
1: 71111 |
1: 71111 |
Лондон |
1: 18 292 |
1: 50 304 |
Ливерпуль, Великобритания |
1: 10215 |
1:112362 |
Манчестер |
1: 7707 |
1: 80925 |
Западная Ирландия |
1 : 7924 |
1: 68670 |
Восточная Ирландия |
1: 5343 |
1: 32 594 |
Бостон, Массачусетс, США |
1: 13914 |
1: 17006 |
Портленд, Орегон, США |
1: 11620 |
1: 33 700 |
Монреаль, Канада |
1: 69442 |
7 |
Окленд, Новая Зеландия |
1: 18168 |
1: 95384 |
Сидней, Австралия |
1: 9818 |
1: 22091 |
Япония |
1:210851 |
1: 70284 |
Израиль (ашкенази) |
1:180000 |
1: 15000 |
Израиль (не-ашкенази) |
1: 8649 |
1: 7111 |
или группы популяций. Полученные данные приведены в табл. 6.4.
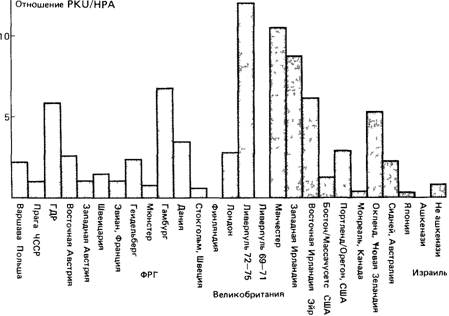
Фенилкетонурия (26160) и гиперфенилаланинемия.
1. Частота ФКУ в восточно-европейских популяциях выше, чем в популяциях запада и югозапада Европы. Различия между популяциями восточной и западной Австрии подчиняются той же закономерности.
2. В скандинавских, и особенно финских, популяциях частота ФКУ исключительно низка. Интересно, что Финляндия отличается от других европейских стран в отношении многих наследственных заболеваний (разд. 6.4.2).
3. В Ирландии ФКУ - довольно распространенное заболевание; различные частоты ФКУ, наблюдаемые в пределах Великобритании, например большое количество случаев в районе Манчестера, возможно, объясняются миграцией из Ирландии.
4. В США частоты ФКУ в Бостоне и Портленде (Орегон) вполне сравнимы с частотами в большинстве популяций Европы. Во франкоязычной области Канады - Монреале ФКУ встречается реже, чем в большей части Европы и двух упомянутых выше городах США, и гораздо реже, чем во Франции, откуда произошла эта популяция.
5. В Японии, единственной стране Дальнего Востока, где проводился скрининг врожденных аномалий, частота ФКУ оказалась особенно низкой, сравнимой только с частотами в популяциях Финляндии и евреев-ашкенази Израиля.
6. Частоты гиперфенилаланинемии (ГФА)- заболевания, не сопровождающегося умственной отсталостью, - изменяются в широких пределах и независимо от частоты ФКУ. Отношение ФКУ/ГФА также варьирует в удивительно широких пределах-от 0,51 до 12,0 (рис. 6.7).
Другие заболевания. В рассматриваемое исследование входят также галактоземия, обусловленная дефицитом трансферазы (23040) (разд. 4.2), гистидинемия (23580), болезнь кленового сиропа (24860) (разд. 4.2.2.4) и гомоцистинурия (23620) (разд. 4.2).
По данным большинства центров скрининга (Гамбурга, Вены, Окленда, Праги, Стокгольма, Цюриха), частоты галактоземии изменялись от 1:30000 и 1:65000. Таким образом, в большинстве популяций европейского происхождения это заболевание встречается со сходными частотами. Частоты в популяциях восточной и западной Австрии значительно отличались друг от друга.
Частота гистидинемии в центрах с наилучшими условиями скринирования варьировала в пределах 1:13 000 – 1:19 000. Гомоцистинурия, лейциноз и (за исключением Монреаля) тирозиноз встречались очень редко, с частотой приблизительно 1:100000 и 1:600000; в Монреале частота тирозиноза достигала 1:13 000, что можно объяснить спецификой франко-канадского изолята.
Для других заболеваний, за исключением фенилкетонурии, скрининг проведен далеко не с такой полнотой, а применяемые методы намного менее надежны. Однако методики улучшают-
6 Популяционная генетика 293
|
Рис. 6.7. Соотношение частот фенилкетонурии (PKU) и гиперфенилаланинемии (НРА) в различных популяциях [1881] |
ся, а с расширением программ скрининга произойдет быстрое увеличение размера выборок Возможно, что некоторые наблюдаемые в настоящее время межпопуляционные различия обусловлены малыми размерами проанализированных выборок, в то время как реальные различия могут пока оставаться невыявленными
Интересно, что ФКУ более распространена в некоторых областях Восточной Германии (ГДР), чем в Западной Германии (ФРГ) Анализ происхождения семей, имеющих детей с ФКУ, показал, что эмигранты из ГДР составляют более высокий процент этих семей, чем ожидается случайно [1761а] Вероятно, частота ФКУ уменьшается с востока на северо-запад Европы Новые сведения относительно истории этих генов скорее всего будут получены в ходе молекулярнобиологических исследований [1363а]
Высокие частоты рецессивных заболеваний в некоторых популяциях Ряд рецессивных заболеваний с высокой частотой встречается в определенных группах популяций Мы уже упоминали тирозиноз в канадских популяциях французского происхождения, другой пример - болезни Тея—Сакса (27280), Ниманна—Пика (25720) и Гоше (23080) в популяции евреев ашкенази. В Финляндии несколько рецессивных заболеваний широко распространились благодаря определенной структуре и истории этой популяции (разд 6 4 2) Некоторые болезни обнаруживаются только в популяциях небольшого размера (изолятах), где их частота высока, а в других популяциях они почти или совсем не встречаются Другие заболевания, такие, как талассемия или серповидноклеточная анемия, имеют высокую частоту только в конкретных географических областях или расовых группах В общем случае рецессивные заболевания, встречающиеся исключительно в определенной популяции малого размера, вызываются генами, не имеющими селективного преимущества в гетерозиготном состоянии, тогда как гены, вызывающие широко распространенные болезни, такие как талассемия и серповидноклеточная анемия, в прошлом имели селективное преимущество у гетерозигот Однако появление метаболически связанных генных мутаций в популяции ашкенази (болезни Тея—Сакса, Ниманна—Пика, Го-
294 6. Популяционная генетика
ше) предполагает наличие в этом случае селективных факторов. Подробно эти вопросы обсуждаются в разд. 6.4.
По мере развития генетического консультирования, распространения пренатальной диагностики, по мере развития методов лечения наследственных заболеваний все большее число стран будет проводить мониторинг наследственных болезней. Выполнение этих программ обогатит наши знания о частоте встречаемости соответствующей патологии.