

6.1. Описание популяций
6.1.1. Закон Харди—Вайнберга: генные частоты
Закон Харди—Вайнберга в случае аутосомных генов [124]. Закон Харди—Вайнберга обсуждался в разд. 3.2. Пусть аллели А1 и А2 имеют частоты А1 = р, А2 = q; р + q = 1; предположим, что скрещивание происходит случайным образом. Тогда частоты фенотипов в популяции будут следующими: p2AiA1, 2pqA1A2, q2A2A2. Это правило можно обобщить следующим образом. Если частоты п аллелей А1, А2, ..., Аn равны p1, p2 …pn и если скрещивание случайно по отношению к данному локусу, то частоты фенотипов определяются случайным попарным сочетанием этих аллелей:
![]()
В отсутствие возмущающих воздействий частоты генов и генотипов остаются постоянными от поколения к поколению. Для аутосомных генов такое состояие «равновесия Харди—Вайнберга» достигается в первом поколении случайного скрещивания. Однако в случае генов, сцепленных с полом, это не так.
Закон Харди—Вайнберга для случая генов, сцепленных с полом. Пусть аллели А1 и А2 встречаются в мужской части популяции с частотами pМ и qM(pМ + qM = 1), тогда фенотипы А1 и А2 будут также иметь частоты рМ и qM. Пусть среди женщин частоты генотипов A1A1, A1A2 и А2А2 равны r, 2s и t(r + 2s + t = 1). Тогда частоты аллелей А1 и А2 у женщин равны рF = г + s и qF = s + t соответственно. Эти женщины образуют ооциты типа A1 и А2 также с частотой pF и qF соответственно. Среди их сыновей аллели А1 и А2 встречаются в том же соотношении. Однако их дочери происходят от яйцеклеток, образовавшихся в результате слияния ооцитов (pFA1 + qFA2) и сперматозоидов, несущих Х-хромосому (рMА1 + + qМА2). Таким образом, следующее поколение будет иметь состав
![]()
Отсюда следует, что частоты аллеля А2 у мужчин и женщин следующего поколения
280 6. Лопуляционная генетика
равны
![]()
Это означает, что а) частота А2 у мужчин каждого поколения равна частоте А2 у женщин предыдущего поколения; б) частота А2 у женщин каждого поколения равна средней частоте этого аллеля у мужчин и женщин предыдущего поколения; в) справедливо равенство
![]()
Следовательно,
если аллельные частоты у мужчин
и женщин в данном поколении не равны,
разность
между ними в следующем поколении
уменьшается вдвое. Кроме того, изменяется
знак этой
разности- если qF
выше
qM,
то
q'M
будет
выше
q'F
Обе аллельные частоты qF
и
qM
сходятся
к общему значению
![]() ,
в
то время как распределение генотипов
у обоих полов стремится к равновесному
состоянию:
,
в
то время как распределение генотипов
у обоих полов стремится к равновесному
состоянию:
![]()
На рис. 6.1 показано, каким образом происходит приближение к состоянию равновесия. Из этого примера следует, что достижение равно ве-
|
Рис. 6.1. Приближение генных частот у мужчин к состоянию равновесия при наследовании, сцепленном с полом, qm = 0,6; qf= 0,3; q = 0,4; сплошной и пунктирной линиями обозначена частота гена у мужчин (qm) и женщин (qf) соответственно, q равновесная частота гена. |
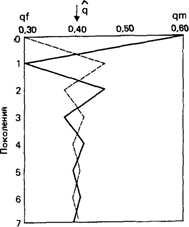
сия Харди Вайнберга после одного поколения случайного скрещивания отнюдь не самоочевидно.
Другие ограничения закона Харди— Вайнберга перечислены в разд. 3.2. Разработке этого фундаментального принципа посвящена значительная часть популяционной генетики.
Генные частоты. Особей, составляющих популяцию, можно классифицировать в соответствии с их фенотипами. В некоторых случаях, особенно в случае генетического полиморфизма, эти фенотипы непосредственно соответствуют генотипам. Однако описание генетической изменчивости популяции на основе частот генотипов оказывается слишком громоздким. Закон Харди —Вайнберга позволяет характеризовать генетическую изменчивость, используя генные частоты. Введение понятия частоты гена упрощает дело; для системы с двумя аллелями вся необходимая информация содержится в одном числе (р или q). Методика определения генных частот в общем виде описана в разд. 3.2 и в приложении 1 (см. также [166; 144]).