

5.1.4.3. Мутации в гемоглобиновых генах и генетический код
Аминокислотные замены в полипептидной цепи отражают соответствующие нуклеотидные замены в ДНК. Так как генетический код известен с начала 1960-х годов, изучение аминокислотных замен, особенно в гемоглобинах, позволило идентифицировать замены оснований в матричной РНК и, поскольку мРНК комплементарна транскрибируемой цепи ДНК, в самих генах. Это можно проиллюстрировать на примере гемоглобина с мутацией серповидноклеточности:
1) изменение биохимического фенотипа: НЬА HbS;
2) аминокислотная замена: глутаминовая кислота валин;
3) кодоны:
Глутаминовая кислота |
Валин |
||
мРНК |
ДНК |
мРНК |
ДНК |
GAA |
стт |
GUA |
CAT |
GAG |
стс |
GUG |
САС |
|
|
GUU |
САА |
|
|
GUC |
GAG |
4) возможные замены оснований в случае, если в ДНК произошла только одна нуклеотидная замена:
СТТ CAT СТС САС;
5) замена основания в ДНК: Т (тимин) А (аденин);
6) типы замены: пуриновое основание замещается пиримидиновым.
Этот тип замены называется «трансверсией». Замена пурина на другой пурин или пиримидина на другой пиримидин известна под названием «транзиции» [1457].
|
Рис. 5.25. Механизм возникновения мутаций на нуклеотидном уровне Возможны четыре транзиции и восемь трансверсий. |
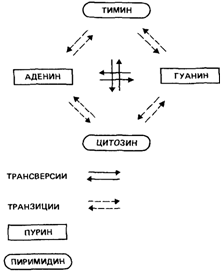
Транзиции происходят чаще, чем можно было бы ожидать, если бы замены оснований носили случайный характер. Любое нуклеотидное основание может заместиться в результате одной транзиции и двух трансверсий (рис. 5.25). Поэтому, если бы направление мутационного процесса было случайным, трансверсии происходили бы вдвое чаще, чем транзиции. Однако из-за вырожденности генетического кода не все нуклеотидные замены приводят к аминокислотным заменам. В табл. 5.16 приведены данные о наблюдавшихся транзициях и трансверсиях различных типов. Транзиции происходит значительно больше, чем можно было бы ожидать в случае, если бы направление мутаций было случайным [1681].
Этот результат нельзя объяснить смещением, обусловленным применением метода электрофореза, выявляющего только те аминокислотные замены, которые приводят к различиям по заряду. В действительности доля «полярных» (и обнаружимых) замен среди всех возможных трансверсий выше, чем среди транзиции. О повышенной частоте транзиции, происходящих в естественных условиях, свидетельствуют также результаты изучения амино-
5. Мутации 191
Таблица 5.16. Замена оснований в разных типах гемоглобина человека [1681] |
|||||||||
|
|
Все замены |
Только те замены, которые приводят к потере функции |
||||||
|
|
α-цепь |
β-цепь |
γ-цепь |
δ-цепь |
Всего |
α-цепь |
β-цепь |
Всего |
Транзиции |
AG |
1 |
10 |
0 |
0 |
11 |
0 |
0 |
0 |
|
GA |
4 |
4 |
0 |
0 |
8 |
0 |
0 |
0 |
|
ТС |
7 |
16 |
4 |
1 |
28 |
7 |
8 |
15 |
|
СТ |
13 |
24 |
3 |
2 |
42 |
9 |
17 |
26 |
Всего |
|
25 |
54 |
7 |
3 |
89 |
16 |
25 |
41 |
Трансверсии |
АТ |
0 |
6 |
0 |
0 |
6 |
0 |
2 |
2 |
|
ТА |
2 |
4 |
0 |
0 |
6 |
2 |
3 |
5 |
|
АС |
3 |
6 |
0 |
0 |
9 |
1 |
1 |
2 |
|
СА |
4 |
5 |
1 |
0 |
10 |
2 |
2 |
4 |
|
GT |
6 |
6 |
1 |
1 |
14 |
6 |
4 |
10 |
|
TG |
4 |
10 |
1 |
0 |
15 |
2 |
6 |
8 |
|
GC |
7 |
5 |
0 |
0 |
12 |
4 |
1 |
5 |
|
CG |
9 |
6 |
0 |
0 |
15 |
6 |
3 |
9 |
Неустановленные трансверсии |
8 |
13 |
0 |
1 |
22 |
0 |
0 |
0 |
|
Всего |
|
43 |
61 |
3 |
2 |
109 |
23 |
22 |
45 |
кислотных замен при дивергенции белков, имеющих общее происхождение, и сравнения родственных форм РНК. Следовательно, вполне вероятно, что при спонтанном мутировании транзиции происходят чаще, чем можно было бы ожидать, если бы направление мутационного процесса было случайным. Этот результат не удивителен, если рассматривать его в свете данных о молекулярных механизмах индуцированных мутаций.
Исследования с использованием многих ферментов рестрикции, которые были предприняты для выявления полиморфных сайтов в уникальной ДНК человека, показали, что с особенно высокой эффективностью такие сайты выявляются с помощью ферментов, опознающих последовательность CpG [1382]. Цитозины в этой последовательности часто метилированы; вместе с тем имеются данные, полученные на других объектах, например, бактериях, свидетельствующие о том, что метилированный цитозин может быть горячей точкой спонтанных мутаций.