

Тип рецептакуліти (receptaculita)
Місце цієї викопної групи тварин в системі безхребетних тривалий час залишалося не визначеним. Вважалося, що вона близька до губок або кишковопорожнинних. За
сучасними уявленнями, ре-цептакуліти — окремий тип, який має своєрідний план будови, перебуваючи на рівні організації губок і археоціат. Викопні бентосні морські організми відомі з ор-•довика до пермського періоду.
Рецептакуліти – кубкоподібні радіальне симетричні організми з вапняковим скелетом, їх розміри становлять від кількох міліметрів до 30—40 см. Скелет складається із зовнішньої та внутрішньої дірчастих стінок. всеоедині є центральна

порожнина, яка ,у верхній частині тварини утворює отвір, можливо, оскулум (рис. 85). Зовні скелет вкритий щільно прилеглими одна до одної ромбоподібними чи шестикутними вапняковими пластинками, розташованими правильними спіральними рядами. Під покривом міститься зовнішня стінка. Вона, як і внутрішня, утворена окремими елементами скелета — меромами. Кожний мером утворений чотирма променями, що розходяться під прямим кутом один до одного в одній площині в зовнішній стінці. Вони з'єднані з такими ж променями внутрішньої стінки за допомогою порожнистої двостінної трубки (радіалі). Кінці променів з'єднані з кІнця'ми променів інших мером і утворюють сітчасті зовнішню та внутрішню стінки. Відомо, що рецептакуліти розмножуються брунькуванням; у молодих особин одразу виникають мероми, зовнішній покрив формується пізніше. Такий спосіб утворення скелета невідомий у інших тварин. Існують кілька десятків видів типу, що об'єднуються в три класи. Одним )3 представників є Receptaculites neptuni.
Тип ортонектиди (orthonectida)
Дрібні (розміром до міліметра) організми, що паразитують у порожнині тіла та статевих залозах морських безхребетних—турбелярій, немертин, поліхет, молюсків, офі-ур. Подібно до пластинчастих і губок тканини та органи в них не розвинені; нервових, м'язових і травних клітин немає. Для них характерне правильне чергування .вільно-живучого (самці та самки) та паразитичного поколінь (рис. 86).
Вільноживуче статеве покоління складається звичайно з самців та самок, хоча відомі й гермафродитні види. У статевих особин тіло вкрите розташованими кільцями клітинами епітелію, частина з яких має війки. Під епітелієм у самок містяться численні яйцеклітини (до 500— 1000), у самців—сім'яник із сперматозоїдами. В епітелії є спеціальний статевий отвір. Навколо сім'яника у самців містяться видовжені скоротливі клітини з мікрофіламен-тами, що тягнуться вздовж усього тіла, а на центральній осі тіла—опорні клітини з товстими волокнами всередині. Видовжені скоротливі клітини самок, розташовані під покривними клітинами.
На передньому кінці тіла в самок і самців є група яе-диференційованих клітин, схожих на паренхіму.
Статеві особини залишають тіло хазяїна. В морській воді вони збираються докупи. Самці виділяють сперму, яка через статевий отвір проникає всередину самки та запліднює її яйцеклітини. Яйця розвиваються всередині самки. В результаті повного та нерівномірного дробіння утворю
ється щільна кулька (морула), що складається з єдиної великої центральної клітини, оточеної дрібними численним ми периферійними клітинами. З морули формується війчаста личинка, яка виходить із материнського організму назовні (див. рис. 86). Личинка вкрита шаром війчастих-
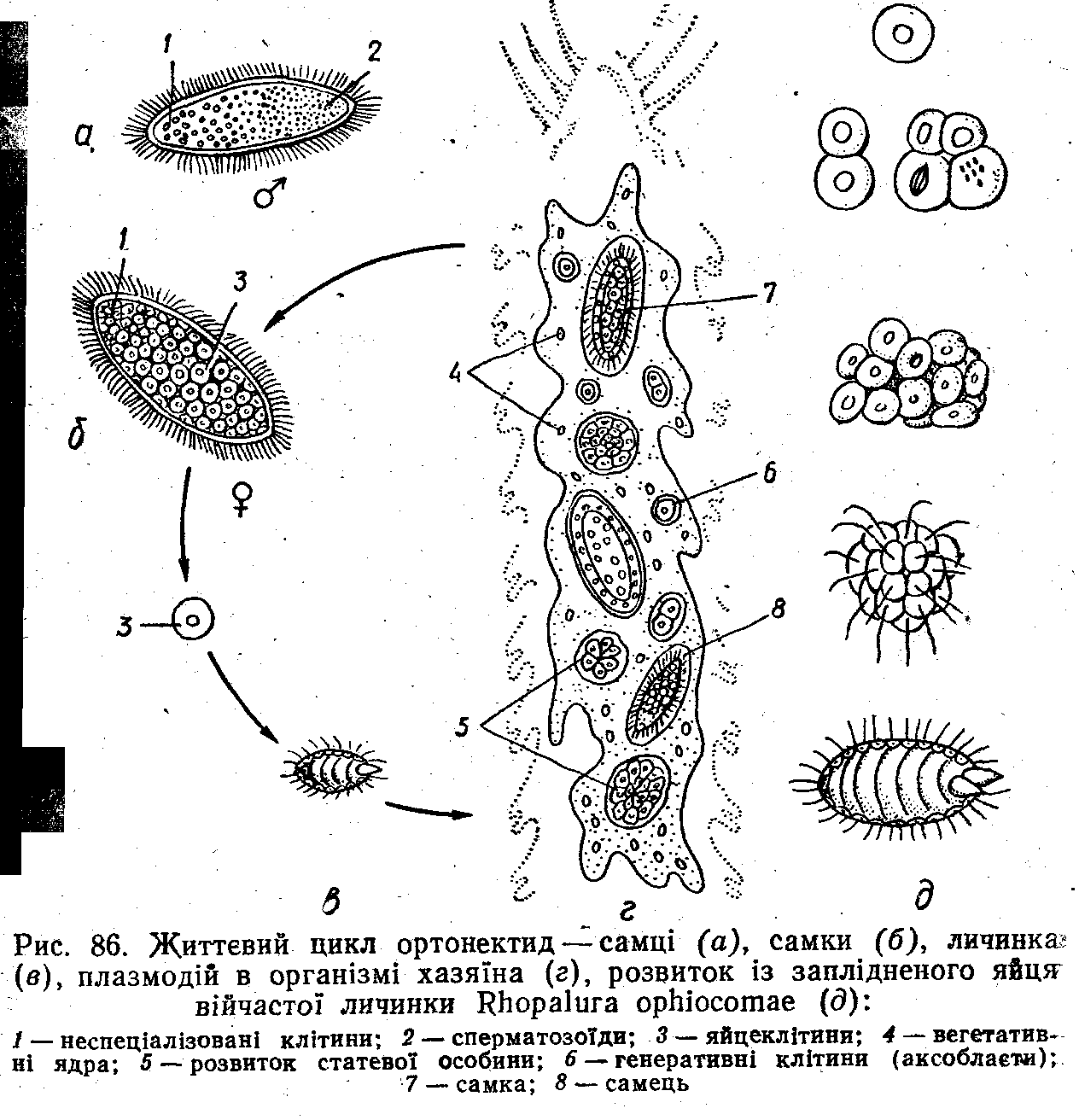
клітин і всередині містить два тільця, що заломлюють світло, та багато зародкових клітин. Личинка деякий час плаває, знаходить хазяїна та проникає в нього, можливо, за допомогою заломлюючих світло тілець.. Усередині хазяїна війчасті клітини гинуть, а зародкові—зливаються в плазмодій.
Паразитичне нестатеве покоління має вигляд багатоядерного плазмодію, що живиться шляхом піноцитозу-та фагоцитозу за допомогою численних виростів, які проникають у тканини хазяїна. Ядрз мітотичне діляться, плаз^ модій росте. Ядра плазмодія поділяються на вегетативні та генеративні. Навколо генеративних клітин відокремлюються ділянки цитоплазми — так виникають генеративні клітини — аксобласти. Одні дослідники вважають розвиток із них статевих особин нестатевим розмноженням, інші — партеногенетичним (статевим, без запліднення). В одних плазмодіях із аксобластів утворюються самці, в інших—самки. Статеві особини виходять у воду (непара-аитичне покоління). „ ч
Представник ортонектид —
паразит офіур R^alfedfa
ophiocomae.
- / (• ,
Місце ортонектид у системі тваринного
царства є предметом дискусій. Одні
вчені вважають їх спрощеними внаслідок
паразитизму нащадками більш
високоорганізова-них багатоклітинних
плоских червів, коловерток, дінофілід,
•ехіурид, інші — однією з груп первинне
примітивних організмів, які залишилися
на рівні організації фагоцители. Найбільш
імовірною є остання точка зору. Оскільки
ор-тонектиди—двошарові тварини,
зовнішній війчастий шар їх тіла можна
прирівняти до кінобласту, внутрішні
клітини — до фагоцитобласту. Вони
перейшли до паразитичного способу
життя у різних групах морських
безхребетних, що спричинило ускладнення
їх життєвого циклу й спеціалізацію
паразитичного покоління.
ТИП ДИЦІЄМІДИ (DICYEMIDA)
До цього типу належать паразити нирок бентосних головоногих молюсків, які досягають 1 см завдовжки. Як і .попередні типи, дицієміди характеризуються відсутністю тканин і органів, рота, кишечника, нервів і м'язів, їх життєвий .цикл дуже складний і до кінця не вивчений. На відміну від попереднього типу, обидва (статеве та нестатеве) покоління диціємід є паразитичними (рис. 87). , У тілі молюсків паразитує ряд послідовних поколінь— гематоген-засновник, кілька поколінь нематогенів, ромбо-ген. Нематоген має витягнуту червоподібну форму. Всередині міститься видовжена осьова клітина, оточена війчастим епітелієм. Вісім-дев'ять передніх епітеліальних клітин утворюють невелике розширення—-головний капор, 14—22 війчасті клітини вкривають тулубний відділ. У цитоплазмі осьової клітини, крім її власного ядра, містяться кілька невеликих ядер, що діляться мітотичним шляхом, утворюючи генеративні клітини, або аксобласти. За їх рахунок всередині осьової клітини материнського нематогена розвиваються нові нематогени, такий процес багаторазово повторюється.
За допомогою електронного мікроскопа на поверхні війчастих клітин знайдено особливі гребінцеві вирости, а між зовнішніми клітинами та осьовою — цитоплазматичні містки (десмосоми). У клітинах диціємід мітохондрії трубчасті, а не пластинчасті, як у більшості багатоклітинних.
Живляться тварини розчиненими речовинами шляхом піноцитозу покривних клітин. Піноцитозні пухирці пере-, даються аксиальній (осьовій) клітині.
Під час утворення нематогена аксобласт спочатку збіль-" шує свій об'єм, потім нерівномірно ділиться мітотичним

шляхом; в результаті утворюється одна велика центральна клітина (макромер), оточена кількома малими (мікро-мерами), що розвиваються у війчастий епітелій. Макромер згодом знову нерівномірно ділиться на велику осьову (аксіальну) клітину та. маленьку клітину, яка проникає всередину великої, ділиться та започатковує аксобласти. Після цих процесів зародок росте за рахунок збільшення розмірів клітин, залишає материнську особину й починає вести самостійне паразитичне життя. Через кілька поколінь нематогенів подібним шляхом з'являються ромбоге-ни. В їх епітелії є особливі бородавчасті клітини з темними зернами екскретів усередині. Більша частина аксобластів усередині ромбогенів дегенерує, а з тих, що залишилися, розвивається нове покоління — інфузоригени.
Інфузориген складається з осьової соматичної клітини, всередині якої виникає кілька безджгутикових сперматозоїдів (сперміїв), та оточуючих її ооцитів. У процесі сперматогенезу та дозрівання ооцитів спостерігається мейоз. Після дозрівання статевих клітин інфузориген розпадається та дегенерує, причому ця стадія існує всередині ромбо-гена. Після запліднення в осьовій клітині ромбогена з зиготи розвивається розселювальна личинка—інфузориформ

(рис. 88). Він укритий війками, всередині має два світло-заломлюючих тільця з гуаніном, а також аксобласти. З сечею молюска інфузориформ виходить назовні, деякий час плаває поблизу дна, а потім інвазує молодого головоно-гого молюска. З аксобластів у тілі молюска .розвиваються ламерівські личинки, які проникають у нирки, де перетворюються на нематогенів-засновників.
Одні дослідники вважають, що аксобласти – це агамети, тоді інфузориформ, нематоген і ромбоген – нестатеві покоління; на думку інших, аксобласти є партеногенетичними яйцями, тоді всі перелічені покоління – партеногенетичні. Остаточно це питання досі не з'ясоване. Не викликає сумнівів лише те, що інфузориген—статеве покоління.
Представник типу – Dicyema. Систематичне положення та походження диціємід так, як і ортонектид, є предметом дискусії вчених, їх вважають спрощеними трематодами або навіть родичами інфузорій. Більш обгрунтованою є думка, що диціеміди, як, і ортонектиди, – це нащадки мечниковської фагоцители.