

Клас коралові поліпи (anthozoa)
Коралові поліпи — найбільш численний клас кишковопорожнинних, до якого належать морські теплолюбні тварини. Описано близько 6 тис. їх видів, із них у Чорному морі знайдено всього чотири, в Азовському—один.
Це поодинокі або здебільшого колоніальні організми. В їх життєвому циклі є лише поліпоїдне покоління, стадія медузи не утворюється. Будова коралових поліпів складніша, ніж гідроїдних. Вони мають ектодермальну глотку, гастральна порожнина в них поділена на камери радіальними перетинками (септами). Замість епітеліально-м'язових є справжні епітеліальні та м'язові клітини. Статеві продукти розвиваються з гастродерми. Більшість. коралів містить вапняковий або роговий скелет.
Тіло окремої особини має форму циліндра (рис. 111). Нижній кінець поодинокого поліпа утворює .пласку підошву, якою він прикріплюється до субстрату, у колоніальних форм він занурений у загальне тіло колонії— ценосарк^ На протилежному кінці міститься ротовий диск, оточений віночком порожнистих щупалець. У одних поліпів (підклас Альціонарії, або Восьмипроменеві—Аісуопагіа, або» Octocorallia) .їх налічується вісім, у інших (підклас Зоан-тарії — Zoantharia) кількість кратна шести або чотирьом. У центрі ротового диска міститься рот, що має форму щілини.
Стінка тіла -коралового поліпа товстіша, ніж у гідроїда,. за рахунок більш розвиненої мезоглеї. Під шаром епідерми, що складається із справжніх епп-еліальних клітин; містяться самостійні поздовжні й кільцеві м'язові клітини, Такі ж клітини є й під шаром гастродерми. Рот веде спочатку в довгу трубку—глотку, .яка вдається .в гастральну порожнину. Глотка утворюється шляхом впинання всередину стінки тіла навколо рота, тому має ектодермальне-походження й вистелена епідермальним епітелієм, що є продовженням зовнішнього покриву. Глотка не цилін-.ідрична, а сплющена, її просвіт має вигляд щілини. У біль-. шості поліпів на одному чи обох боках цієї щілини розташовані ротові жолобки — сифоногліфи, клітини яких мають дуже довгі війки. За допомогою цих війок всередину гастральної порожнини постійно надходить свіжа вода,. а у поліпів, які живляться найдрібнішими планктонними організмами, разом з нею й їжа. З гастральної порожнини вода з неперетравленими рештками їжі виводиться назовні через іншу частину глотки. Таким чином відбувається дихання тварин, а в деяких — і живлення.
Гастральна порожнина в
коралових поліпів не суцільна, а
поділена на камери -перетинками, або
септами, якї відходять від внутрішньої
поверхні стінки тіла. У верхній частині
поліпа септи зростаються зі стінками
глотки, в нижній — вільно звисають у
гастральну порожнину, залишаючи
центральну її частину — «шлунок» —
неподіленою. Септи побудовані з двох
шарів гастродерми з мезоглеєю між ними.
Крім того, в кожній септі є поздовжній
м'язовий валок. Клітини гастродерми
.в різних ділянках сепг неоднакові.
Більша їх частина, як і внутрішня стінка
тіла» складається з джгутикових
епітеліальних клітин, які виконують
переважно транспортну функцію, але
здатні й до
фагоцитозу. Вільні
краї, септ, що вдаються в г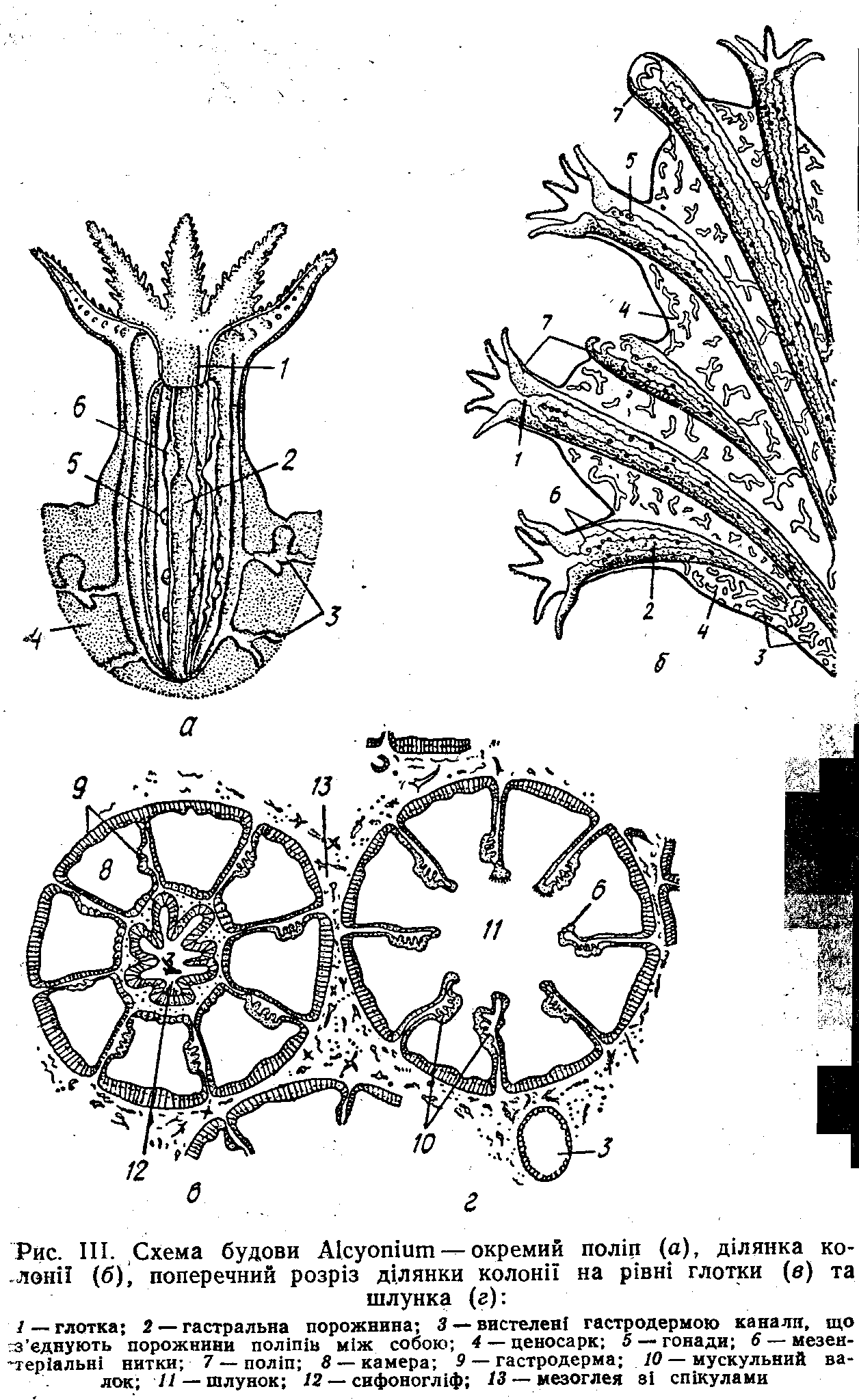 астральну
порожнину, утворюють потовщення —
мезентеріальні нитки,
які складаються із залозистих і
фагоцитуючих клітин. У багатьох видів
у нижній частині поліпа вони продовжуються
в довгі тонкі внутрішні щупальця,
аконції,
на яких, крім зазначених типів клітин,
є щ& й жалкі клітини. У деяких видів
аконції викидаються через рот або
спеціальні отвори й виконують функції
захисту та травлення (рис, 112).
астральну
порожнину, утворюють потовщення —
мезентеріальні нитки,
які складаються із залозистих і
фагоцитуючих клітин. У багатьох видів
у нижній частині поліпа вони продовжуються
в довгі тонкі внутрішні щупальця,
аконції,
на яких, крім зазначених типів клітин,
є щ& й жалкі клітини. У деяких видів
аконції викидаються через рот або
спеціальні отвори й виконують функції
захисту та травлення (рис, 112).
Вивчення механізмів травлення актиній свідчить, що здобич, яку тварина проковтує, через глотку потрапляє в центральну частину гастральної порожнини — «шлунок». Мезентеріальні нитки змикаються навколо їжі, аконції обплутують її, утворюється тимчасово замкнена порожнина, де й відбувається травлення. Перша фаза травлення проходить в. «шлунку» під дією травних ферментів, що виділяються залозистими клітинами, а друга—в фагоцитуючих клітинах епітелію мезентеріальних ниток. Після цього краї септ розходяться, між ними з'являються щілини, й поживні речовини, що утворилися в результаті травлення, розносяться за допомогою джгутикового епітелію стінок септ на периферію гастральної порожнини. Отже, в коралів, як і у сцифоїдних медуз, має місце диференціювання гастродерми на травний відділ (клітини мезентеріальних ниток) і транспортний (стінки сент і внутрішні стінки тіла поліпа).
Коралові поліпи живляться всіма способами, характерними для сидячих донних безхребетних. Серед коралів є активні фільтратори й високоспеціалізовані хижаки, яким властиві унікальні механізми захоплення та проковтування здобичі. Багато з них поєднують обидва типи живлення. Крім того, всі, вони поглинають розчинені у воді органічні речовини клітинами епідерми за рахунок спеціальних механізмів активного транспорту. І, нарешті, вони здатні до живлення за рахунок симбіотичних водоростей.
Нервова система коралових поліпів, як і гідроїдних, має вигляд дифузного плексуса. Органів чуття на відміну від вільноплаваючих медуз у них немає. Проте корали реагують на хімічні подразнення, що виявляється у їх вибірковому ставленні до їстівних і неїстівних речовин. Шматочки їжі проковтуються актинією, а індиферентні предмети (наприклад, скляні кульки) відкидаються. Очевидно, поодинокі чутливі клітини реагують на хімічні речовини їжі.
Лише деякі групи коралових
поліпів, наприклад актинії, не мають
скелета. Восьмипроменеві коралові
поліпи (підклас
Alcyonaria) мають внутрішній скелет, який
міститься в мезоглеї. Вапняковий
скелет складається з окремих
мікроскопічних тілець — склеритів
або голок — спікул,
які утворюються всередині особливих
клітин—склеробластів.
У деяких видів спікул так багато, що
вони зливаються, утворюючи суцільне
мереживо (див. рис. 111). Роговий скелет
скріплює окремі спікули або утворює
 суцільний
стрижень у мезоглеї. Хоча скелет у
восьмипро-меневих поліпів міститься в
мезоглеї, за походженням він ектодермальний,
оскільки склеробласти мігрують із
епідерми до мезоглеї, там розмножуються
й продукують скелетні елементи.
суцільний
стрижень у мезоглеї. Хоча скелет у
восьмипро-меневих поліпів міститься в
мезоглеї, за походженням він ектодермальний,
оскільки склеробласти мігрують із
епідерми до мезоглеї, там розмножуються
й продукують скелетні елементи.
У склерактиній (підклас Zoantharia ряд Scleractinia) скелет зовнішній, вапняковий; він утворюється клітинами епідерми та має вигляд чашечки (склеротеки) навколо кожного поліпа. Від склеротеки всередину тіла вростають скелетні перетинки (склеросепти), які проходять всередині м'яких септ, що підрозділяють гастральну порожнину на камери. Склеросепти глибоко впинають стінки тіла поліпа в його гастральну порожнину (див. рис. 118). Скелет у
склерактиній суцільний. У багатьох видів чашечки сусідніх поліпів зливаються, утворюючи масивний скелет із отворами, де розташовані окремі особини. У деяких видів (наприклад, мозковик) поліпи зливаються докупи, і вся колонія набуває вигляду масивного утворення в формі півкулі, по поверхні якої проходять звивисті борозни, що надає коралу вигляду мозку. У центрі борозен проходить суцільна щілина, утворена із злитих ротів поліпів; з її боків рядами розташовані численні щупальця. У мозкови-ка неможливо встановити межі окремих поліпів—вся колонія е єдиним «надорганізмом» (див. рис. 119).
Коралові поліпи розмножуються нестатевим і статевим шляхами. Нестатеве розмноження відбувається різними способами. Найпоширеніші — брунькування, поперечний чи поздовжній поділ окремих поліпів, лацерація, що призводять до збільшення кількості особин. Крім того, колоніальні склерактинії здатні до розмноження, подібного до вегетативного розмноження рослин — фрагментація колонії та приживлення її уламків; вихід окремих голих поліпів із екзоскелета в товщу води з наступним їх осіданням, за рахунок чого починає розвиватися нова колонія; від-брунькування окремих поліпів із наступним їх відокремленням і утворенням нових колоній; просторовий розділ колоній; відокремлення кінців гілок колоній схоже на автотомію (самокалічення).
Найчастіше в розгалужених колоній (наприклад, Асго-рога) під час шторму або внаслідок порушення цілісності свердлячими організмами (губками, молюсками тощо) відбувається їх фрагментація. В результаті утворюються уламки колоній, які містять живі 'поліпи. Такі фрагменти переміщуються під дією прибою та течій, врешті-решт потрапляють на твердий субстрат і приживаються.
У склерактиній з родин Caryophyllidae і DendrophyHi-<іае утворюються бруньки, з'єднані з краєм материнської колонії вузькою перетинкою; такі бруньки періодично відпадають і, потрапляючи на придатний субстрат, започатковують нові колонії.
За несприятливих умов (висихання, надмірне або недостатнє освітлення) у деяких склерактиній (наприклад, Seriatopora histris) окремі поліпи ізолюються від ценосар-ка, випадають із своїх скелетних чашечок у воду і розносяться течіями. На новому місці вони знаходять придатний субстрат, осідають і започатковують нові колонії.
Під час статевого розмноження в гастродермі септ утворюються гонади. Корали можуть бути роздільностатевими або гермафродитами. Статеві продукти коралів через рот викидаються назовні; запліднення відбувається у воді. Із заплідненого яйця виходить личинка'— плинула. Личинка вкрита війками, має рот (на вегетатив^ ному полюсі) та гастральну порожнину. На протилежному (анімальному) полюсі міститься група чутливих клітин із довгими війками. Така личинка плаває, живиться дрібними планктонними організмами, потім опускається на субстрат анімальним полюсом (на цьому місці утворюється підошва) й перетворюється на поліпа. У деяких ви-, дів виходить більш пізня личинка, що нагадує поліпа та має щупальця (актинула). Колонії утворюються шляхом брунькування первинного поліпа.
Клас Anthozoa поділяється на два підкласи: Альціонарії (Аlсуоnаrіа) та Зоантарії (Zoantharia).
Підклас Альціокарії, або Восьмипроменеві корали (Alcyonaria, або Octocorallia)
Це колоніальні корали. Поліп має вісім пірчастих щупалець і вісім септ у гастральній порожнині. Його внутрішній скелет залягає в мезоглеї. Підклас поділяється на багато рядів, з яких найважливішими є Альціонацеї (Аlсуоnасеа), Рогові корали, або Горгонацеї (Gorgonacea), Морські пера (Pennatulacea) та Столоніфери (Stolonifera).
Ряд Alcyonacea - Альціонацеї
Це м'які корали, скелет яких складається з дрібних розкиданих у мезоглеї вапнякових голок — спікул. Центральний (осьовий) скелет у них ніколи не утворюється,

тому їх колонії стеляться або мають вигляд куль і грибів, які широкою основою прикріплюються до субстрату (рис. 113). Альціонацеї—найбагатший у видовому відношенні ряд восьмипроменевих коралів, вони поширені в морях і океанах, проте найбільша кількість видів мешкає на мілководді тропічних морів.
У більшості альціонацей сильно розвинена мезоглея, тому колонії мають масивний вигляд. До цього ряду належить широковідомий Alcyonium digitatium, колонія якого 'нагадує кисть руки, та яскраво забарвлені деревоподібне розгалужені колонії Gersemia fruticosa.
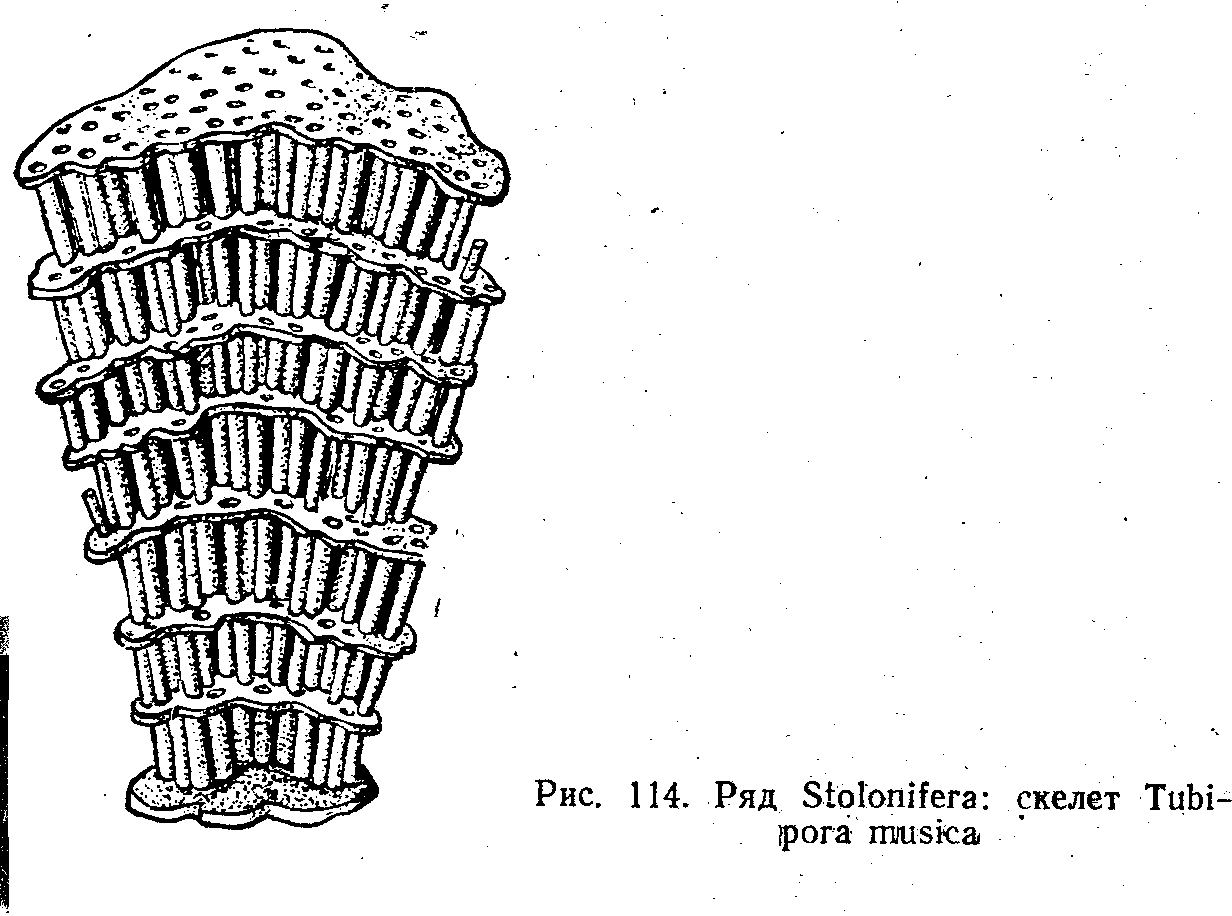
Ряд Столоніфери (Stolonifera)
Це корали, в яких скелетні спікули в мезоглеї зливаються й утворюють трубочки, зв'язані між собою поперечними пластинками. У живих колоніях у трубочках містяться гастральні порожнини поліпів.
У тропічному поясі часто-трапляється «органчик» — Тubiрога musica (рис. 114), колонії якого нагадують музичний інструмент — орган. У розправленому стані поліпи, які виступають з трубочок, мають яскраво-зелене забарвлення. Коли вони скорочуються і втягуються у трубочки, колонія набуває малинового кольору, в який забарвлений скелет.
Ряд Горгонарії, або Рогові корали (Gorgonacea)
До цього ряду належать корали, які утворюють розгалужені деревоподібні або бичоподібні гнучкі колонії. Для горгонарій характерна наявність щільного осьового скелета, який пронизує стовбур і гілки колонії (рис. 115). Він складається зі з'єднаних вапном або роговою речовиною спікул чи рогових пластинок. Крім осьового скелета, в мезоглеї містяться окремі спікули у вигляді голочок; паличок, веретен, густо вкритих горбками, відростками, лусочками, тощо.
К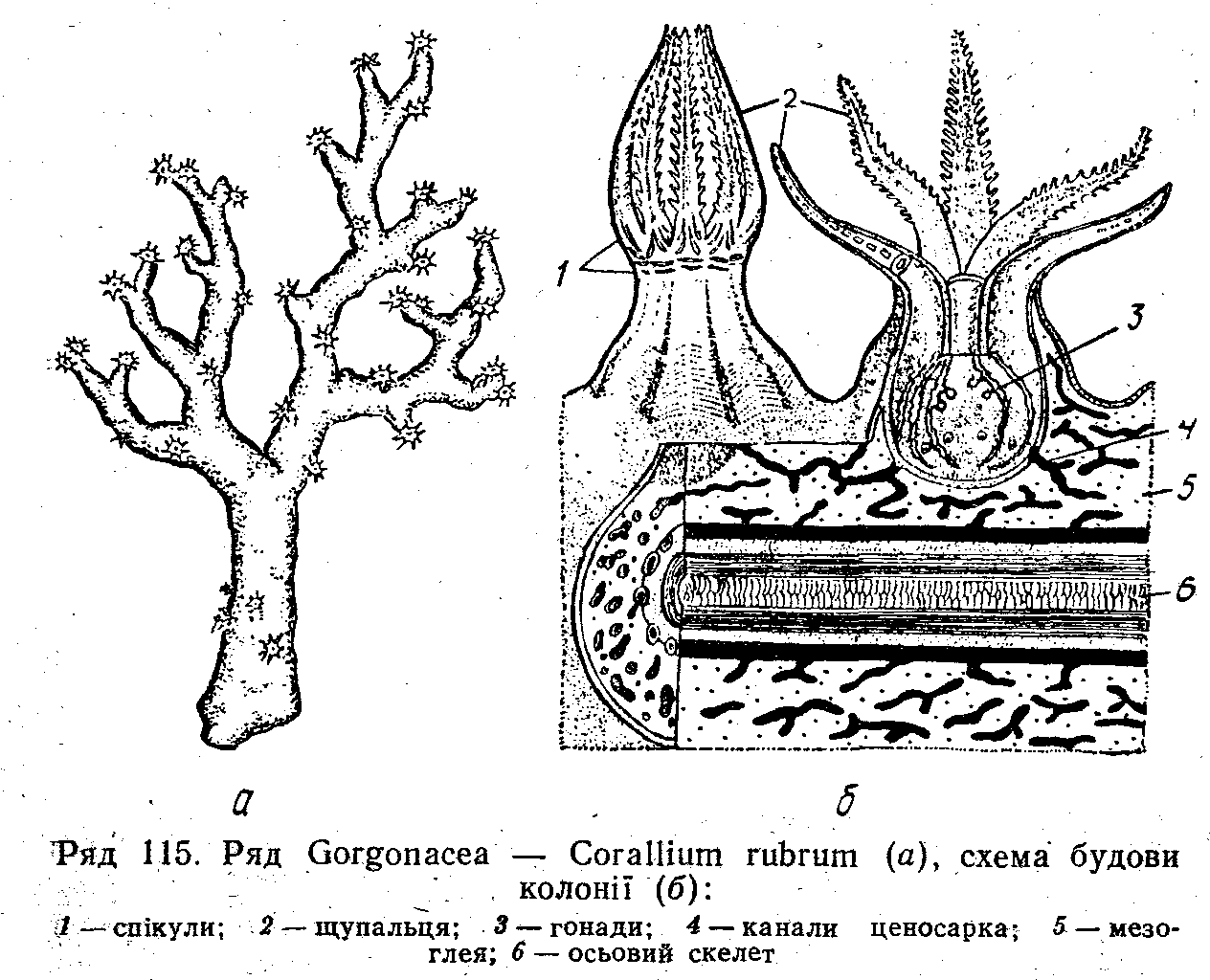 олонії
горгонарій міцно прикріплюються до
твердого субстрату розширеною основою.
Це переважно мілководді тварини,
найбільше їх мешкає в тропіках. Серед
них
олонії
горгонарій міцно прикріплюються до
твердого субстрату розширеною основою.
Це переважно мілководді тварини,
найбільше їх мешкає в тропіках. Серед
них
відомий червоний, або благородний, корал (Corallium rubrum), твердий скелет якого використовується для виготовлення прикрас.
Осьовий скелет більшості видів горгонарій складається з рогової речовини, просякнутої вапном, він дуже міцний і разом з тим пружний. Наприклад, різні види Gorgonia мають вигляд широкої ажурної пластини до 2 м завдовжки та до 1,5 м завширшки. Вони оселяються в зоні прибою, але їх роговий скелет має таку гнучкість, що велетенські віяла витримують могутні удари хвиль. Ці витончені колонії надзвичайно яскраво забарвлені. Разом з іншими горгонаріями вони утворюють на. невеликій глибині ділі підводні «ліси», забарвлені в жовті, червоні, коричневі, фіолетові кольори.