

Часть V Основные типы экосистем и их динамика
Дидактическая единица 5
Тема 11: динамика и устойчивость экосистем
Методические рекомендации
Тема «Динамика и устойчивость экосистем» позволяет провести предварительное обобщение и повторение материала значительной части курса «Общая экология».
Дидактические материалы раздела «Основные типы экосистем» приводятся в учебном пособии «Введение в экологию», книга 1 (Глазачев С.Н., Косоножкин В.И.).
ВАША ОЦЕНКА по 100-балльной шкале (дидактическая единица 5)
Вид работы |
Оценка, баллы |
Подпись преподавателя |
|
Максимум |
Реальная |
||
1. Работа на практических занятиях по теме 11 |
25 |
|
|
2. Домашнее задание по теме 11 |
20 |
|
|
3. Контрольная работа и тестирование |
55 |
|
|
ИТОГО по дидактической единице 5 |
100 |
|
|
Таблица 5.
Календарно-тематический план аудиторных занятий по дидактической единице 5
Тема занятий |
Лекции |
Практические |
||
Кол-во часов |
Дата |
Кол-во часов |
Дата |
|
1. Динамика и устойчивость экосистем |
2 |
|
3 |
|
2. Текущий контроль (контрольная работа) |
- |
|
1 |
|
ИТОГО: |
2 |
|
4 |
|
Глава XI Динамика и устойчивость экосистем (тема 11)
11.1. Динамика и устойчивость экосистем (конспект лекции и хрестоматия)
Устойчивость природных экосистем
Биогеоценоз (элементарная экологическая система) по определению В.Н. Сукачёва – «<...> внутренне противоречивое, диалектическое единство, находящееся в постоянном движении и развитии». Постоянство экосистем относительно, они непрерывно изменяются. Мы говорим о стабильности, постоянстве экосистемы при отсутствии значительных, принципиальных изменений, причём за короткий (в геологическом масштабе) отрезок времени.
Устойчивость природных экосистем (по Н.В. Дылису, 1978):
- Под устойчивостью биогеоценозов (элементарных экосистем) следует понимать сохранение структурно-функциональной организации их в неизменном или мало измененном состоянии в течение достаточно продолжительного времени. Это свойство природных биогеоценозов есть результат длительного эволюционного процесса. В ходе его, с одной стороны, совершенствовались сами организмы, с другой – шла взаимная подгонка организмов и популяций друг к другу и к косной среде (упрощенно: в результате естественного отбора сохранялись не только самые совершенные организмы, но и самые эффективные биоценозы и экосистемы).
- В результате современные природные биогеоценозы приобрели известный запас прочности и с помощью отрицательных обратных связей выработали способность сохранять тип своей структуры и ритмы работы даже при значительных изменениях внешних условий.
- В силу исторически сформировавшейся прочности организации современные природные биогеоценозы оказываются способными к многократному восстановлению по присущему «унаследованному» ими типу организации при разного рода разрушительных воздействиях со стороны внешних факторов. Например, на месте распаханной ковыльной степи при прекращении использования постепенно через ряд стадий восстанавливается исходный тип ковыльной степи.
- Устойчивость экологических систем не имеет, однако, абсолютного характера; систем, всегда себе равных, не изменяемых, не существует. Устойчивость их относительна и ограничена определенными отрезками времени, разными для разных биогеоценозов.
- Наиболее устойчивые биогеоценозы – природные, занимающие в пространстве возвышенные местообитания, на которых они развились естественным путем. Их современная организация лучше других подогнана к зональным климатическим условиям, и они наиболее автономны в своем функционировании. Такие наиболее устойчивые биогеоценозы (экосистемы) получили название климаксовых.
Ю. Одум (1986) подчёркивает информационную, кибернетическую (от греч. kybernetike – искусство управления) природу устойчивости естественных экосистем: «Помимо потоков энергии и круговоротов веществ... экосистемы характеризуются развитыми информационными сетями, включающими потоки физических и химических сигналов, связывающих все части системы и управляющих ею как одним целым». Всё многообразие гомеостатических механизмов в экосистемах он подразделяет на два основных типа:
1) механизмы, основанные на существовании в системе отрицательной обратной связи;
2) механизмы, связанные с избыточностью функциональных элементов в системе.
Отрицательные обратные связи стабилизируют экосистему: «При внешнем воздействии на систему, находящуюся в динамическом равновесии, система реагирует таким образом, чтобы ослабить эффект внешнего воздействия» – принцип Ле Шателье.
Наряду с действием механизмов обратной связи, устойчивость экосистемы может обеспечиваться избыточностью функциональных элементов или экологическим дублированием (Одум, 1986). Например, если фотосинтез в экосистеме осуществляют три группы растений, имеющих различные температурные оптимумы, то суммарная интенсивность фотосинтеза на выходе может оставаться относительно постоянной даже при изменении температуры на входе (рис 11.1).
Рис. 11.1. Поддержание стабильности в экосистеме при избыточности функциональных элементов (по Одуму, 1986 с изменениями, пояснения в тексте).
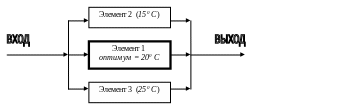
Исходя из принципа избыточности, можно предположить, что устойчивость экосистем будет возрастать с увеличением видового разнообразия их биоценозов. Во многих случаях это утверждение справедливо, однако специальные исследования показывают, что в определенных (часто экстремальных) условиях окружающей среды наиболее устойчивы те сообщества, которые включают небольшое число специализированных видов организмов.
Безусловно, определяющую роль в процессах стабилизации и регуляции экосистемы и окружающей среды играет биотическое сообщество (биоценоз). В ходе эволюции различные виды организмов приобрели следующие хорошо вам известные способности:
1) адаптации к изменению параметров окружающей среды;
2) целенаправленного изменения и регулирования окружающей абиотической среды (образование почвы, поддержание постоянства рН, концентраций биогенов и параметров микроклимата);
3) авторегуляции плотности численности популяции;
4) образования надорганизменных биологических систем (симбиотических мутуалистических систем, консорций, сообществ и биоценозов);
5) выполнения специализированной работы по поддержанию системы (разделение на продуцентов, консументов и редуцентов, формирование неперекрывающихся экологических ниш у доминирующих видов, дифференциация «стратегий жизни» (виолентов, эксплерентов, патиентов, передвигающихся и непередвигающихся организмов), а также многие другие особенности, позволяющие осуществлять процессы авторегуляции.
Циклические и поступательные изменения экосистем
Различают два основных типа изменений экосистем во времени: циклические изменения и поступательные изменения.
Циклические изменения экосистем отражают периодичность повторения внешних условий (суточную, сезонную, многолетнюю). Такие изменения повторяются регулярно, и к ним адаптированы члены биоценоза. Экосистема подмосковного ельника-черничника зимой разительно отличается от летнего состояния, но это различные аспекты одной и той же экосистемы, которые мы наблюдаем регулярно. В геоботанике (фитоценологии) регулярно повторяющиеся сезонные изменения в составе и облике фитоценоза принято называть аспектами. Этот термин часто используют и в экологии – говорят о весеннем или зимнем аспекте экосистемы (биогеоценоза).
Напомним, что закономерные циклические колебания интенсивности и характера биологических процессов и явлений называются биологическими ритмами (см. тему 2). Экологическое значение биоритмов состоит в том, что они дают возможность живым организмам приспосабливаться к циклическим изменениям абиотических условий окружающей среды (смена дня и ночи, сезонов года, приливов и отливов), а также координировать процессы жизнедеятельности особей одного вида и разных видов (периоды размножения, наличие пищи).
Поступательные изменения экологических систем, в отличие от циклических, не исчезают при возобновлении внешних условий. Эти изменения могут быть более или менее значительными, накапливаться постепенно или происходить относительно быстро, обуславливаться преимущественно внутренними (по отношению к экосистеме) или внешними причинами. Развитие экологических систем, приводящее к значительному изменению видового состава их биоценозов и всех экологических взаимодействий, носит название экологической сукцессии.
Экологическая сукцессия (от лат. successio – преемственность) – последовательная смена во времени одних биоценозов другими на определённом участке земной поверхности. Классический пример экологической сукцессии – это многолетний путь развития экосистемы от первых поселенцев, осваивающих обнажения горных пород, через ряд промежуточных стадий до зрелой экосистемы, находящейся в динамическом равновесии с окружающей средой.
Основоположником теории сукцессии растительных сообществ был американский ботаник Клементс Фредерик Эдвард (Clements F.E.) (1874 – 1945). О сукцессии растительных сообществ он писал:
«Так как растительность развивается, то одна и та же площадь начинает последовательно заниматься разными растительными сообществами. Этот процесс называется растительной сукцессией. В пределах одного региона результатом серий этих последовательных стадий является одна и та же финальная, или климаксовая, стадия, начинаются ли они от водной поверхности или твёрдой скалы, или обнажённой земли ...» (Клементс, 1929; цит. по Кузнецовой).
«Развитие климаксовой формации (зрелого сообщества) состоит из нескольких существенных процессов ... 1) обнажение (участка суши), 2) миграция, 3) (приживание), 4) конкуренция, 5) взаимодействие, 6) стабилизация... Они последовательны на начальных стадиях и взаимодействуют наиболее сложным образом – на всех последних» (Клементс, 1928; цит. по Кузнецовой).
Типы экологических сукцессий
Принято различать первичные и вторичные сукцессии
Первичные сукцессии начинаются на субстратах, практически не затронутых процессами почвообразования (вулканическая лава, обнажения горных пород, пески после отступления воды, отложения аллювия). В ходе первичной экологической сукцессии организмы, взаимодействуя между собой и с окружающей средой, в прямом и переносном смысле слова сами готовят почву для следующего этапа сукцессии, поэтому первичные сукцессии занимают многие десятки и даже сотни лет. Смена этапов в ходе экологической сукцессии протекает закономерно – в направлении от простых и неустойчивых сообществ к более сложным и устойчивым.
Вторичные сукцессии протекают в более благоприятных для организмов условиях, так как в этом случае полностью или частично сохраняется почва. Классический пример вторичной сукцессии – зарастание заброшенных пахотных земель (залежей). В степных условиях сукцессия на залежи проходит довольно быстро: 1) стадия однолетних сорняков; 2) стадия «крупного бурьяна», преимущественно полыней; 3) стадия корневищных злаков с примесью бобовых растений; 4) стадия дерновинных злаков. На 12 (25) год после прекращения вспашки появляются ковыли и восстанавливается существовавшее до начала вспашки сообщество. Вторичные сукцессии, приводящие к постепенному восстановлению существовавших на этом участке экосистем, называют демутационными (восстановительными) сукцессиями.
Приведём некоторые термины и определения, используемые для описания сукцессии. В ходе сукцессии принято выделять несколько характерных этапов (фаз). Эти этапы образуют так называемую сукцессионную серию, а каждый характерный этап носит название сериальной стадии или просто стадии. Первые этапы сукцессии обычно называют пионерными стадиями. На последней стадии сукцессии формируется устойчивое, зрелое, климаксовое сообщество или климакс.
Климакс (от греч. klimax – лестница) – заключительная стадия экологической сукцессии, наиболее стабильное состояние экосистемы, находящейся в равновесии с окружающей средой.
Закономерные изменения экосистемы в ходе сукцессии завершаются формированием наиболее устойчивой системы (климакса), которая находится в состоянии динамического равновесия с окружающей средой и теоретически может существовать неопределённо долго при отсутствии внешних катастрофических воздействий.
Идея целостного подхода к развитию биотического сообщества принадлежит Фредерику Клементсу, который считал, что биотическое сообщество (биоценоз) является сверхорганизмом. В монографии «Растительная сукцессия» (1916 г.) Клементс подчёркивал, что биотическое сообщество (биоценоз) в своём развитии, подобно любому живому организму, проходит ряд последовательных и закономерных стадий вплоть до состояния зрелости (климакса). Следовательно, все биотические сообщества (биоценозы) развиваются в одном направлении и в итоге приходят к одному и тому же результату – климаксовому сообществу, причём для данных климатических условий существует только один вариант климакса (это так называемая теория моноклимакса).
Теории моноклимакса обычно противопоставляют теорию поликлимакса, согласно которой, в данных климатических условиях могут существовать несколько типов устойчивых климаксовых экосистем, соответствующих мозаике различных местообитаний. Климаксовые сообщества на водораздельных территориях соответствуют понятию климатический климакс, а на склоновых почвах или почвах, сформировавшихся на контрастных почвообразующих породах, образуются своеобразные эдафические климаксы.
В.Н. Сукачёв (1964) отрицал наличие некоторой завершающей стадии в развитии биогеоценозов (экосистем). Он писал: «Необходимо отметить, что учение о климаксовой растительности могло возникнуть лишь при недооценке того большого влияния, которое оказывают растительный и животный мир, а также микроорганизмы на другие компоненты биогеоценоза. Можно лишь говорить о том, что биогеоценотический процесс с течением времени замедляется, а растительный покров и вместе с ним и животный мир могут приобретать некоторую относительную устойчивость, но и только. Поэтому если и пользоваться термином «климакс», то только в смысле приобретения растительностью или биогеоценозом относительно большей замедленности в развитии ...»
Аутогенные и аллогенные (экзогенные) сукцессии
Аутогенная сукцессия. Если сукцессионные изменения определяются главным образом внутренними (по отношению к экосистеме) причинами, такими как накопление мертвых растительных остатков, продуктов метаболизма, конкуренцией, деятельностью землероев, то их называют аутогенными (самопорожденными) сукцессиями.
Обычно выделяют две группы причин аутогенных сукцессий:
а) изменения, связанные с взаимодействием популяций разных видов в биоценозе (прежде всего, конкуренцией). Такие изменения В.Н. Сукачёв называл сингенетическими (взаимное сживание, «притирка» различных видов организмов, происходящая, как правило, на ранних стадиях формирования биоценоза);
б) изменения, связанные с преобразованием абиотической среды в результате саморазвития биогеоценоза (экосистемы). Такие изменения В.Н. Сукачёв называл эндодинамическими (эндогенетическими). В процессе жизнедеятельности организмы постоянно и непрерывно изменяют среду своего обитания. Постепенно накапливаясь, такие изменения приводят к нарушению сложившегося в экосистеме равновесия, что в свою очередь, ведёт к изменению видового состава биоценоза.
Аллогенная (экзогенная) сукцессия. Если сукцессионные изменения определяются, главным образом, внешними причинами – то это аллогенные (экзогенные, порожденные извне) сукцессии.
Н.В. Дылис (1978) подразделяет аллогенные сукцессии на следующие типы:
а) климатогенные, обусловленные общими изменениями теплового режима или атмосферного увлажнения и захватывающие изменения экологических систем на обширных территориях;
б) геоморфогенные, связанные с динамикой рельефа в результате тектонических движений земной коры, развития долин, оврагов и т. д.;
в) почвенно-грунтовые, связанные с изменением уровня грунтовых вод или размывом почвенного слоя атмосферными осадками и другими процессами;
г) зоогенные и фитогенные, связанные с внедрением в сложившиеся биоценозы ранее отсутствовавших там видов, обладающих высокой конкурентоспособностью.
Деление экологических сукцессий на аллогенные (экзогенные) и аутогенные весьма проблематично, поскольку в реальных условиях все факторы действуют на систему совокупно, и сложно выделить одну главную (основную) причину наблюдаемой экологической сукцессии.
Деятельность человека, связанная с нерациональным использованием окружающей среды, очень часто приводит к возникновению экзогенных (аллогенных) сукцессий даже в стабильных природных экосистемах. Причем наступающие изменения идут в направлении обеднения видового состава биоценозов, деградации почв, снижения устойчивости экосистем. Сукцессии такого типа принято называть дигрессиями.
Учитывая масштабы воздействия человека на биосферу, можно утверждать, что в современных условиях аутогенные сукцессии в чистом виде практически отсутствуют или, по крайней мере, протекают на фоне постоянных аллогенных возмущений, вызываемых антропогенными факторами.
Примеры экологических сукцессий
Пример первичной сукцессии на скалах, в лесах Урала (Чернова, Былова 2004):
1-ая стадия. «Поселение накипных лишайников, которые сплошь покрывают каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток и нематод. Мелкие клещи-сапрофаги и первичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, в основном после смачивания скал дождём или туманом.
2-ая стадия. Преобладание листоватых лишайников, постепенно образующих сплошной ковер. Под круговинками лишайников в результате действия выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идёт отмирание слоевищ и накопление детрита. Под лишайниками в большом количестве обитают мелкие членистоногие: коллемболы, панцирные клещи ... и другие, образуется микрогоризонт, состоящий из их экскрементов.
3-тья стадия. Поселение литофильных мхов. Они погребают лишайники и подлишайниковые плёночные почвы. Ризоиды мхов прикрепляются не к камню, а к мелкозёму, мощность которого достигает уже 3 см. Под мхами колебания температуры и влажности уже значительно меньше, чем под лишайниками. Усиливается деятельность микроорганизмов и увеличивается разнообразие групп животных.
4-ая стадия. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растёт участие более крупных беспозвоночных-сапрофагов: энхитреид, дождевых червей, личинок насекомых».
Здесь приводится описание только четырех этапов первичной экологической сукцессии. Обратите внимание на то, что взаимодействуя между собой и с окружающей средой, организмы на каждом этапе в прямом и переносном смысле слова сами готовят почву для следующего этапа сукцессии. Причем смена этапов в ходе экологической сукцессии протекает закономерно – в направлении от простых и неустойчивых систем к сложным и устойчивым системам.
Другой классический пример первичной сукцессии – песчаные дюны в штате Индиана США у южной оконечности озера Мичиган (по Ю. Одуму, 1986):
«... Отступая к своим современным границам, озеро оставляло все более и более молодые песчаные дюны. Поскольку субстрат песчаный, сукцессия протекает медленно, и можно наблюдать серию сообществ разного возраста: пионерные стадии у берегов и все более старые стадии по мере удаления от берега. Это своеобразная «природная лаборатория сукцессий».
Первые поселенцы на дюнах – это прибрежные злаки, ива, песчаная вишня, хлопковое дерево и животные, такие как бегающие по песку длинноногие жуки-скакуны, живущие в норах пауки, кузнечики. Вслед за сообществом первых поселенцев следуют открытые сухие леса из сосны Банкса, затем чёрного дуба и, наконец, на самых старых дюнах – влажные леса из дуба и гикори или бука и клена.
Несмотря на то, что развитие сообщества начинается в очень сухом и бесплодном местообитании, оно в конечном итоге приводит к влажному и холодному сомкнутому лесу, резко отличающемуся от голых дюн. Мощная, богатая гумусом почва, населенная дождевыми червями, контрастирует с сухим песком, на котором она образовалась. Использовав современные методы радиоуглеродного датирования, Олсон установил, что для достижения климаксового лесного сообщества на дюнах озера Мичиган требуется около 1000 лет».
Классический пример вторичной сукцессии – зарастание заброшенных пахотных земель (залежей). В степных условиях сукцессия на залежи проходит довольно быстро:
1. Стадия однолетних сорняков.
2. Стадия «крупного бурьяна», преимущественно полыней.
3. Стадия корневищных злаков с примесью бобовых растений.
4. Стадия дерновинных злаков.
На 12 (25) год после прекращения вспашки появляются ковыли – восстанавливается существовавшее до начала вспашки сообщество.
Вторичные сукцессии, приводящие к постепенному восстановлению существовавших на этом участке экосистем, называют демутационными (восстановительными) сукцессиями.
«Формирование экосистемы – динамический процесс. Её формируют виды, дополняющие друг друга и соответствующие другим видам и условиям среды.
На пригодном для жизни субстрате биотические сообщества последовательно сменяют друг друга, формируют закономерный ряд экосистем, ведущий к наиболее устойчивой в данных условиях климаксовой экосистеме. Смена фаз (стадий) сукцессии идёт в соответствии с определёнными правилами. Каждая фаза (стадия) готовит условия для возникновения последующей фазы.
Когда система приближается к состоянию климакса, в ней, как и во всех равновесных системах, происходит замедление всех процессов развития. Например, восстановительная сукцессия темнохвойного леса в южной тайге может идти следующим образом. Сначала быстро меняется ряд лугово-кустарничковых фаз с образованием берёзового или осинового леса. Затем может произойти замещение этого леса сосной, либо прямое вытеснение елью. Сосну относительно быстро вытесняет ель или пихтово-кедровый лес. Эти климаксовые системы отнюдь не лишены тенденций к развитию. Вековая сукцессия идет по типу заболачивание – разболачивание или остепнение – расстепнение.
Эмпирический закон сукцессионного замедления есть следствие закона Г. Одума и Р. Пинкертона: аутогенная сукцессия идёт в направлении фундаментального сдвига потока энергии в сторону увеличения её количества, направленного на поддержание системы. С этим законом связано и правило стабилизации видового состава климаксового сообщества: видовое разнообразие стремится к пику на ранних или средних стадиях сукцессии, а затем снижается и стабилизируется в климаксе. В ходе сукцессии происходит увеличение замкнутости биогеохимических круговоротов веществ. Хотя все круговороты не полностью замкнуты, но степень незамкнутости варьирует в очень широких пределах» (Реймерс, 1994).
Ещё один пример – сукцессия природного заболоченного озера: «Заболоченные озёра – это маленькие водоёмы, которые развиваются в понижениях рельефа в районах бывшего оледенения с холодным влажным климатом северных лесов. ... Наиболее отличительную черту заболоченных озёр составляет ковёр растительности – сплавина, плавающая на воде и качающаяся под ногами. Сплавина нарастает в направлении к центру озера, и открытая водная поверхность постепенно, в течение столетий или даже тысячелетий после окончания ледникового периода, сокращается. По истечении достаточно длительного периода времени озеро заполняется органическим веществом и превращается в «сухую» территорию. ... Ниже даётся описание типичной болотной сукцессии для территории северо-центральной части США:
1. Открытая водная поверхность хорошо развитых заболачивающихся озёр окрашена в кофейный цвет из-за растворённого в воде органического вещества, или гумуса, поступающего от сплавины. ... Животный и растительный мир в болотной воде довольно беден.
2. Вокруг открытой водной поверхности расположена сплавина, сформированная осоками и болотным мхом. ... Сплавина нарастает и захватывает поверхность водоёма, одновременно увеличивая свою толщину и занимаемую ею площадь.
3. Утолщение сплавины ведёт к появлению кустарников... Многие из светолюбивых растений сплавины, затеняемые кустарниками, исчезают, торф сплавины, связанный корнями, становится более плотным и теперь уже не колышется при ходьбе по нему.
4. В кустарниковую стадию внедряются деревья нескольких видов, среди них одно из наиболее характерных – лиственница. Под пологом деревьев большинство кустарников погибает. ...
5. Эти первые деревья со временем замещаются другими. Вместо лиственницы появляются ель или бук и клён, которые господствуют в большинстве лесов этого региона.
Болотная сплавина развивается под влиянием свойственных ей внутренних процессов: произрастания растений, увеличения массы торфа за счёт корней и отмерших остатков, замещения низкорослых растений более высокими и мощными. ... Сплавина со временем смыкается со дном озера, и слой торфа становится сплошным, что позволяет сформироваться здесь устойчивому лесу» (Р. Уиттекер, 1980).
Олиготрофное озеро – устойчивая экосистема. Но в результате избыточного поступления в водоём биогенных элементов (из-за сброса неочищенных сточных вод или распашки территории прилегающего водосбора) может начаться бурное развитие планктона, накопление донных отложений, интенсивное развитие прибрежной растительности и др. В конечном итоге чаша водоёма заполнится донными отложениями, и водная экосистема прекратит своё существование.
Закономерности изменения экосистем в ходе аутогенной сукцессии
По мнению Ю. Одума (1986), представление о том, что экологическая сукцессия – это закономерный процесс развития, а не просто случайная смена функционирующих изолированно видов, остается одной из самых важных объединяющих теорий в экологии. С другой стороны, знание основных закономерностей экологической сукцессии позволит человеку управлять её ходом.
I. Тенденции изменения экосистем в ходе аутогенной сукцессии (по Р. Уиттекеру, 1980):
1. «В ходе сукцессии обычно происходит прогрессивное развитие почвы, которое сопровождается возрастанием её мощности, увеличением содержания органического вещества и дифференциацией почвенных горизонтов в направлении к зрелой почве климаксового сообщества.
2. Увеличиваются высота, биомасса и ярусное расчленение растительных сообществ.
3. Запас элементов минерального питания, содержащихся в почве и растениях, возрастает, и возрастающая доля его содержится в тканях растений.
4. Продуктивность (скорость формирования органического вещества на единицу площади) возрастает одновременно с развитием почвы и структуры сообщества и усилением использования сообществом ресурсов среды.
5. По мере увеличения высоты и сомкнутости надземной части сообщества микроклимат экосистемы все более определяется характером самого сообщества.
6. Видовое разнообразие увеличивается в направлении от простых сообществ ранних стадий сукцессии до богатых видами более поздних стадий или до зрелого сообщества.
7. Значимость отдельных популяций попеременно возрастает и падает, и они сменяют одна другую вдоль градиента времени подобно тому, как это происходит в устойчивых сообществах по градиентам условий среды.
Скорость замещения в ходе сукцессии одних видов другими во многих случаях замедляется по мере смены небольших по размерам недолговечных растений более крупными и более долговечными.
8. Относительная устойчивость сообщества возрастает. Ранние стадии в некоторых случаях явно неустойчивы, имеют популяции, быстро сменяющие одна другую; сообщество конечной стадии обычно стабильно, с доминированием долгоживущих растений, которые поддерживают свои популяции в составе сообщества, неизменяющимся больше в определенном направлении».
II. Тенденции изменения экосистем в ходе аутогенной сукцессии (по Ю. Одуму, 1986, с изменениями)
Энергетика экосистемы
1. Возрастают биомасса (В) и количество органического детрита
2. Возрастает валовая первичная продукция (Рg) за счёт чистой первичной продукции; вторичная продукция изменяется мало
3. Увеличивается дыхание (R) всего сообщества (R = Ra + Rh)
4. Уменьшается чистая продукция (Р) всего сообщества (P = Pg – R)
5. Соотношение Pg/R приближается к единице (закон Одума – Пинкертона)
6. Соотношение В/Рg возрастает
Круговорот биогенных элементов
7. Круговороты элементов становятся всё более замкнутыми
8. Увеличиваются время оборота и запас важнейших элементов
9. Удерживается и сохраняется больше биогенных элементов
Виды и структура сообщества
10. Меняется видовой состав сообщества (флористические и фаунистические эстафеты)
11.** r-стратеги замещаются К-стратегами
12. Усложняются и удлиняются жизненные циклы
13. Увеличивается величина организмов и (или) их стадий расселения (семян, молоди)
14. В значительной степени развивается взаимовыгодный симбиоз
Стабильность
15. Возрастает резистентная устойчивость
16. Снижается упругая устойчивость
Общая стратегия
17. Возрастает эффективность использования энергии и биогенных элементов
** Уточнение терминов и определений
r-стратеги – виды с высоким потенциалом размножения, но низкой способностью к выживанию в условиях конкуренции;
К-стратеги, наоборот, виды с относительно низким потенциалом размножения, но высокой конкурентоспособностью.
Резистентная устойчивость – способность противостоять внешним воздействиям.
Упругая устойчивость – способность быстро восстанавливаться после прекращения внешнего воздействия.
Если на первых стадиях развития экосистемы первичная продукция заметно превосходит расходы на дыхание сообщества (накапливается биомасса и мёртвая органика), то такую сукцессию называют автотрофной.
Гетеротрофной называют сукцессию, которая начинается в среде, обогащенной органическим веществом, поступившим извне или накопленным предыдущим сообществом. В этом случае (например, после попадания в пруд больших количеств органики) на первых стадиях сукцессии расходы на дыхание сообщества значительно превышают первичную продукцию самой экосистемы. Современную цивилизацию можно сравнить с затянувшейся стадией гетеротрофной сукцессии – человечество активно расходует запасы органики (уголь, нефть, газ), накопленные в предыдущие геологические эпохи.
Закон Г. Одума – Р. Пинкертона
Закон Одума-Пинкертона (1955) – в ходе аутогенной сукцессии возрастает валовая первичная продуктивность (ВПП), но увеличивается и количество энергии, направленной на поддержание экосистемы, расходы на дыхание всего биотического сообщества (R), по мере того, как накапливаются биомасса и органическое вещество. В зрелых, климаксовых экосистемах отношение ВПП/R приближается к единице. Экспериментальные данные, подтверждающие закон Одума-Пинкертона, приведены в табл. 11.1.
Таблица 11.1. Годовая продукция и дыхание (в ккал/м2 в год) в развивающихся и устойчивых экосистемах, (по Ю. Одуму, 1986 г).
Экосистемы |
ВПП |
Ra |
ЧПП |
Rh |
ЧПП – Rh |
Поле люцерны |
24400 |
9200 |
15200 |
800 |
14400 |
Молодые посадки сосен |
12200 |
4700 |
7500 |
4600 |
2900 |
Дубово-сосновый лес среднего возраста |
11500 |
6400 |
5000 |
3000 |
2000 |
Большой ручей |
20800 |
12000 |
8800 |
6800 |
2000 |
Спелый дождевой тропический лес |
45000 |
32000 |
13000 |
13000 |
0 |
Зрелый лес умеренной зоны |
8000 |
4600 |
3400 |
3200 |
200 |
Прибрежный пролив |
5700 |
3200 |
2500 |
2500 |
0 |
Открытый океан |
1000 |
400 |
600 |
600 |
0 |
Пустыни |
200 |
100 |
100 |
100 |
0 |
Пруд, после сброса сточных вод |
2000 |
900 |
1100 |
9000 |
– 7900 |
Здесь: ВПП – валовая первичная продуктивность, Ra – автотрофное дыхание, ЧПП – чистая первичная продуктивность, Rh – гетеротрофное дыхание, (ЧПП – Rh) – чистая продукция сообщества (в ккал/м2 в год).