

3. Эмоции, эмоциональная память, эмоциогенные структуры мозга
Систематическое изучение эмоций было начато ещё Ч. Дарвином, опубликовавшим в 1872 году обстоятельный труд «Выражение эмоций у человека и животных». Уже в этой работе отмечалась связь эмоций с мышечной активностью, то есть интеграция в феномене эмоций телесных изменений с психическими. Экспериментальное исследование механизмов эмоций и локализация эмоциогенных структур нервной системы были начаты в 20-х годах нашего столетия У. Кенноном. Кеннон, в частности, пришёл к выводу о связи эмоционального поведения с таламическими образованиями мозга. (Таламус – зрительный бугор – одно из наиболее крупных образований древней коры). Этим было положено начало представления о центральных механизмах эмоций.
В настоящее время психическим и телесным проявлениям и механизмам эмоций человека и животных посвящено большое число исследований, однако содержание самого термина «эмоции» всё ещё не вполне определилось; разные авторы вкладывают в него несколько различный смысл. Е.А. Громова [21] суммирует те существенные моменты, которые являются общими для наиболее полных определений эмоций, следующим образом: «Этим общим является, во-первых, выражение субъективного переживания человеком собственного состояния, обусловленного воздействием окружающего мира. Иначе говоря, эмоции выступают в качестве своего рода аппарата, оценивающего эти воздействия со знаком плюс или минус, на что неоднократно указывал П.К. Анохин [53]. Во-вторых, … указание на существенные «телесные изменения» … двигательную активность и изменение вегетативных процессов». В соответствии с наиболее общепринятой в настоящее время активационной теорией эмоций (Линдсли [54, 55]) основной функцией эмоций является изменение уровня активации мыслительной деятельности в зависимости от значимости и новизны ситуации. Наиболее детально и систематически активация мыслительной деятельности посредством вовлечения в неё механизмов эмоций исследована применительно к феномену эмоциональной памяти [20, 21, 56-65, 88, 92-102]. Под эмоциональной памятью понимается «способность пережитое ранее эмоциональное состояние в комплексе с воспоминанием о вызвавшей его ситуации и субъективным отношением к ней. Эмоциональная память обладает своими отличительными чертами, главными из которых являются: быстрота формирования, большая прочность и непроизвольность воспроизведения» [57]. Изучение механизмов эмоциональной памяти привело к выявлению определяющей роли в формировании эмоций нейрональных структур, в которых проведение нервного импульса осуществляется моноаминами: норадреналином (норадренергические структуры) и серотонином (серотонинергические структуры).
Вопрос о структурах, формирующих эмоции и управляющих ими, вообще говоря, не является вполне однозначным. Выделение тех или иных структур, как эмоциогенных, различными исследователями в определённой мере зависит от того содержания, которое вкладывается в понятие «эмоции». Следуя активационной концепции эмоций [54, 55, 56], мы должны считать эмоциогенными те структуры мозга, которые управляют уровнем активации коры головного мозга, уровнем его реактивности на изменения, происходящие во внешнем мире. Назовём основные структуры, оказывающие активирующее воздействие на нейроны головного мозга.
Ретикулярная формация ствола головного мозга была описана венгерским анатомом И. Ленхошшеком ещё в 1855 году. Интерес исследователей к этой структуре в последние десятилетия резко возрос в связи с открытием в 1949 году её активирующей роли Мэгуном и Моруцци. Они пробуждали спящее животное, стимулируя ретикулярную систему через электроды, вживлённые в ствол мозга. Филогенетически ретикулярная формация – одна из наиболее древних нейрональных структур мозга, она играет важную роль в регуляции уровня активности центральной нервной системы у всех позвоночных [50, 67]. Её нейроны вмести с их дендритами и аксонами образуют сложную пространственную сетку, что и определяет название структуры (от латинского reticulum – сеточка).
У человека ретикулярная формация расположена выше спинного мозга в продолговатом мозгу и тянется до промежуточного мозга. Одной из особенностей ретикулярной системы является то, что в ней отсутствует чёткая структурная организация: в таком протяжённом образовании нейроны кажутся расположенными довольно случайным образом.
Ретикулярная формация связана со всеми органами чувств. Сенсорные импульсы, вызванные световыми, звуковыми или осязательными ощущениями, поступают не только к специализированным нейронам мозга, но и к ретикулярной формации через отростки – коллатерали, которые отдают этой структуре все сенсорные пути, проходя через ствол мозга. К ретикулярной системе ствола мозга поступают также импульсы от проприоцепторов (рецепторов, которые находятся в мышцах, суставных сумках и вестибулярном аппарате внутреннего уха), контролирующих положение тела и тонус скелетной мускулатуры, а также от коры головного мозга. Кроме того, имеет место импульсация ретикулярной системы от внутренних органов, которая не связана с конкретными ощущениями и воспринимается, как «общее состояние» («тёмные чувства», описанные И.М.Сеченовым еще в 1866 году).
Все эти воздействия, обусловленные информацией, поступающей в мозг от органов чувств, в частности, информацией, относящейся к напряжению мускулатуры и двигательной активности, а также степенью интенсивности мыслительной деятельности коры головного мозга и состоянием организма определяют уровень активации ретикулярной формации, которая, в свою очередь, через нервные волокна восходящей
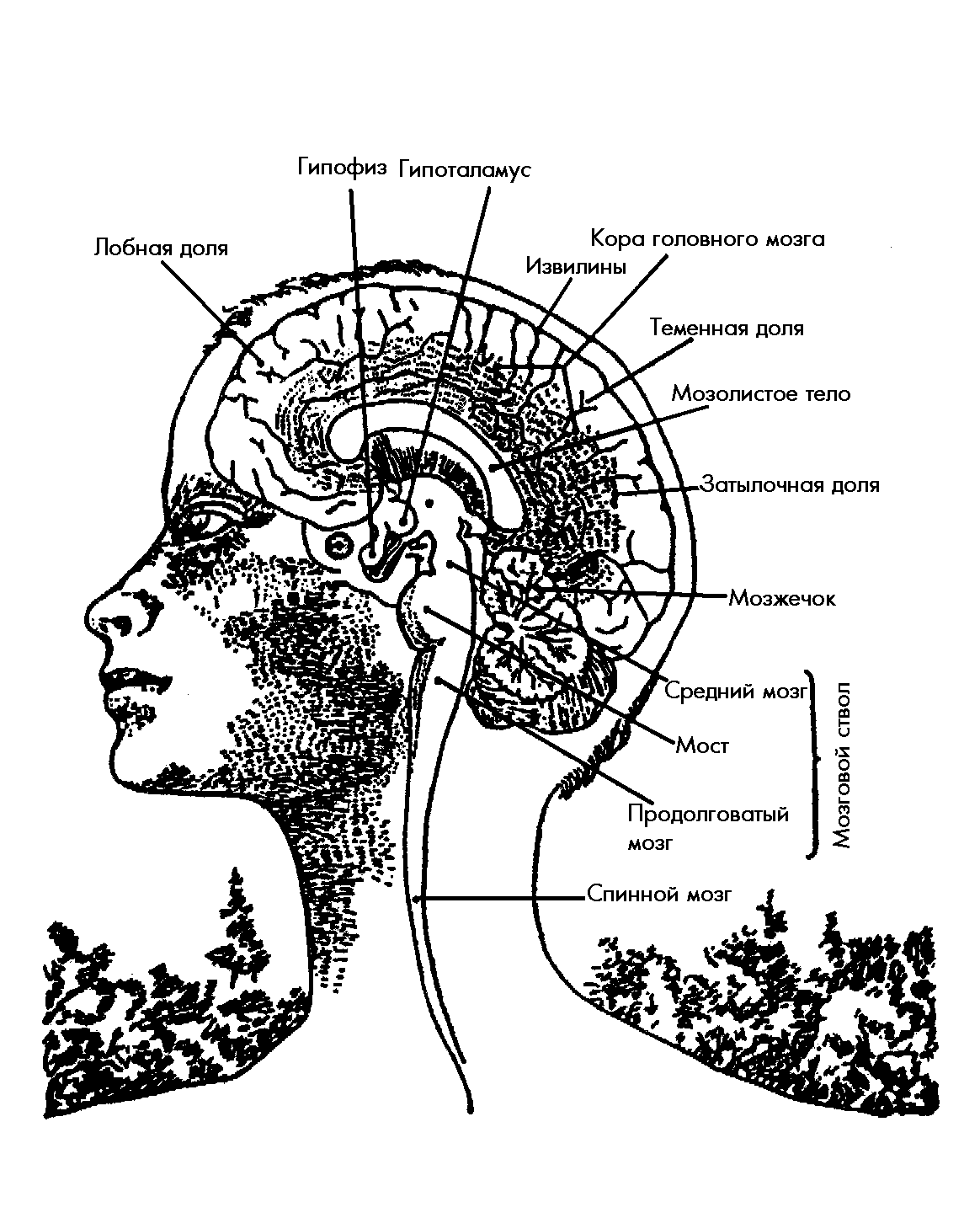
Рис.2. Схематическое изображение мозга, показывающее взаимное расположение важнейших структур центральной нервной системы.
активизирующей системы (Мэгун и Моруцци, 1949) поддерживает необходимый общий тонус («уровень алертности» [68]) коры больших полушарий. Таким образом, степень ретикулярной активности полностью определяется импульсами, поступающими в ретикулярную формацию. Она лишена собственной активности и является общей «системой тревоги» [50], которая отвечает одинаково на все приходящие к ней импульсы. Её функция – обеспечить общее «пробуждение» всей массы нейронов обоих полушарий. Поддержание уровня активности осуществляется, по-видимому, корой головного мозга, хотя и через посредство ретикулярной формации.
Отметим двухсторонний характер взаимодействия коры головного мозга и ретикулярной формации. Такая кольцеобразно замкнутая (а не однонаправленная) система включает обратную связь, что обеспечивает возможность саморегулировки в системе кора–ретикулярная формация.
Ретикулярная формация, обладая двусторонними – афферентными и эфферентными – связями со спинным мозгом (её эфферентные пути спускаются почти до поясничного отдела спинного мозга), влияет на мышечный тонус, на реакции, связанные с изменением позы, и другие виды рефлекторной деятельности спинного мозга. В частности, активность ретикулярной формации находится в связи с ориентировочными рефлексом, описанным И.П. Павловым ещё в 1910 году. Внезапное изменение в окружающей животное среде вызывает соответствующие изменения в его поведении. Так, в ответ на неожиданный звуковой сигнал собака вздрагивает, резко поднимает голову, поворачивается в сторону звука. Описанные в предыдущем разделе внешние проявления «Ага-переживания» весьма сходны с проявлениями ориентационного рефлекса с той лишь разницей, что в изменении позы отсутствует компонента, связанная с поиском источника сигнала. Сигнал – зарождающаяся новая идея – возникает в сознании самого человека. Можно сказать, что «Ага-переживание» есть ориентировочный рефлекс, направленный внутрь.
Ретикулярная формация регулирует и ряд вегетативных функций. В её нижних структурах, расположенных в продолговатом мозге, имеются участки, раздражение которых вызывает изменение дыхания, сосудистого тонуса и сердечной деятельности, кашель и даже рвоту. Хорошо известны такие вегетативные сдвиги, связанные с эмоциональными состояниями, как учащение дыхания и сердцебиения, или изменение сосудистого тонуса, проявляющегося в том, что человек бледнеет или, напротив, краснеет, взволнованное покашливание, появление пота, в особенности, на ладонях и ступнях ног. С активностью ретикулярной формации многие исследователи связывают и КГР.
В значительной степени активизирующее воздействие ретикулярной формации осуществляется не непосредственно, а при участии гипоталамуса, с которым эта структура имеет двухсторонние связи.
Гипоталамус (подбугорье) представляет небольшое (вес около 5 граммов) образование, расположенное непосредственно под таламусом, рис.2, или зрительным бугром (отсюда его название) и образующее дно третьего желудочка (одна из полостей головного мозга, заполненная внутримозговой жидкостью-ликвором). Это одна из наиболее сложных структур головного мозга; гипоталамус существует у всех хордовых, что указывает на его жизненно важное значение. Гипоталамус образован специализированными группами клеток – ядрами, которые имеют широкие связи как друг с другом, так и с другими структурами центральной нервной системы. В частности, гипоталамус имеет мощные двухсторонние связи с корой головного мозга, в особенности с её ассоциативными лобными долями, с ретикулярной формацией и со спинным мозгом (его вегетативными центрами) [49, 50, 69-74].
Важная особенность анатомического строения гипоталамуса – высокая проницаемость его сосудов (в особенности в области ядер заднего гипоталамуса) для многих биологически активных веществ, в частности, для моноаминов и некоторых высокомолекулярных белковых соединений. Вследствие этого гипоталамус находится в условиях, резко отличающихся от тех, в которых функционируют другие структуры центральной нервной системы, защищённой от многих веществ, циркулирующих в кровяном русле, гематоэнцефалическим барьером [49]. Гипоталамус – единственная структура мозга, которая может оказывать гуморальные (то есть через посредство кровотока) воздействия на организм и получать информацию о состоянии гуморальной среды организма.
Гуморальные влияния на организм гипоталамуса осуществляют, главным образом, мелкоклеточные ядра заднего гипоталамуса, которые в ответ на приходящие к ним нервные импульсы секретируют в кровеносные сосуды, обеспечивающие кровоснабжение гипофиза, биологически весьма активные высокомолекулярные белковые соединения – рилизинг-факторы, которые управляют весьма известными функциями аденогипофиза (передняя часть гипофиза). На воздействие рилизинг-факторов различные группы клеток гипофиза отвечают синтезом и секрецией в кровяное русло гормонов, осуществляющих центральную регуляцию всей эндокринной системы. Между гипоталамусом и гипофизом существует взаимосвязь, что даёт основание объединять обе структуры в единую гипоталамо-гипофизарную систему. Детальное описание этой системы будет дано ниже в связи с её участием в формировании длительных адаптационных реакций. Здесь отметим лишь интересную деталь: ответвление третьего желудочка проникает в гипофиз, поэтому возможна связь гипофиза с гипоталамусом и рядом других нейрональных структур, расположенных у стенок третьего и четвёртого желудочков и соединяющей их узкой полостью сильвиева водопровода, через внутримозговую жидкость.
Гипоталамус является также структурой, обеспечивающей высшее центральное представительство вегетативной нервной системы. Вегетативные эффекты, возникающие при раздражении различных участков гипоталамуса, имеют различную направленность. Швейцарский физиолог В. Гесс ещё в 1925 году предложил разделять физиологические реакции на две группы: эрготропные и трофотропные. Эрготропными были названы системы, регулирующие мобилизацию и расходование энергетических ресурсов клетки, органов или организма в целом для активации и поддержания их деятельности, трофотропными – системы, регулирующие восстановление и накопление энергетических ресурсов, процессы анаболизма, ассимиляции [49]. Задние ядра гипоталамуса, по Гессу, относятся преимущественно к эрготропной системе головного мозга. Раздражение этих ядер сопровождается хорошо известными проявлениями активации симпатической нервной системы: расширением зрачков, учащением сердцебиения, повышением артериального давления, расширением сосудов скелетной мускулатуры и, вследствие этого, повышением её тонуса, угнетением перистальтики кишечника.
Медиатором, осуществляющим передачу нервных импульсов в симпатических образованиях, является норадреналин. Норадреналин вместе с его предшественником в метаболических процессах дофамином и адреналином относится к числу катехоламинов – классу биологически активных веществ, образующихся в организме из аминокислот путём цепочки последовательных превращений. Помимо прямого медиаторного действия, норадреналин, поступая, подобно гормонам в кровь (а также в тканевую жидкость), принимает активное участие в регуляции многих функций.
Катехоламины, содержащиеся в крови, оказывают сильное влияние на протекание и результат всех процессов в организме, вызывая эффект того же характера, что и при возбуждении симпатической нервной системы. Исключительную роль играют катехоламины в осуществлении функции симпатической нервной системы, названной академиком Л.А. Орбели адаптационно-трофической. Значение катехоламинов, как регуляторов экстренных приспособительных механизмов, следует из их способности оказывать быстрое и мощное влияние на процессы метаболизма, стимулировать активность ферментов, осуществляющих распад гликогена и жиров, повышать содержание глюкозы в крови, способствовать окислению жирных кислот, повышать потребление кислорода тканями, увеличивать работоспособность сердца и скелетной мускулатуры, обеспечивать общее перераспределение крови для интенсивного снабжения тканей энергетическими ресурсами, усиливать возбуждение центральной нервной системы (в первую очередь, её катехоламинэргических – норадрен- и дофаминэргических структур), участвовать в формировании эмоциональных реакций [49].
Передние отделы гипоталамуса, согласно классификации В. Гесса, относятся к трофотропной системе головного мозга. Раздражение этих отделов сопровождается преимущественно парасимпатическими эффектами: сужением зрачков, снижением артериального давления, повышением тонуса и перистальтики пищеварительного тракта, расслаблением скелетных мышц и так далее. Нейромедиатором парасимпатической нервной системы и соответствующих ядер гипоталамуса является ацетилхолин – сложный эфир холина и уксусной кислоты. До недавнего времени было общепринято представление, что ацетилхолин выполняет исключительно медиаторную функцию. В настоящее время результаты исследований, выполненных, главным образом, во Всесоюзном научно-исследовательском институте физической культуры, позволяют заключить, что ацетилхолин поступает из органов и тканей в кровь и активно участвует в гуморальной регуляции функций, стимулируя протекание многих трофотропных процессов. Избыточный ацетилхолин крови связывают эритроциты, которые разносят его по организму. При соответствующих локальных изменениях в органах, тканях или жидких средах организма он может высвобождаться и принимает участие в трофотропных процессах на различных уровнях, вплоть до клеточного [49].
Однако вернёмся к гипоталамусу. «Можно, сильно схематизируя, сказать, что задние (эрготропные) передние (трофотропные) отделы гипоталамуса интегрируют соответственно функции симпатической и парасимпатической вегетативной нервной системы и находятся в антагонистических отношениях между собой. Первые в основном обеспечивают непосредственную подготовку и создают условия для активной деятельности организма во внешней среде, вторые по преимуществу обеспечивают отдых и восстановление сил» [74].
Итак, гипоталамус оказывает широкое активирующее воздействие на центральную нервную систему и на весь организм. Это воздействие осуществляется тремя путями: 1) непосредственно через обширные нервные связи с другими областями головного мозга, 2) через вегетативную нервную систему, 3) гуморальным путём, главным образом, через посредство гипофиза. Выше отмечалась ещё одна возможность воздействия системы гипоталамус–гипофиз на другие структуры мозга – через внутримозговую жидкость.
До сих пор мы описывали структуры, которые можно отнести к эмоциогенным по двум признакам. Эти структуры участвуют: 1) в активации мыслительной деятельности, 2) в формировании внешних проявлений эмоций. Однако ещё со времён Аристотеля принято считать, что для эмоций характерна ещё одна черта: они, как отмечалось выше, оценивают события внешней и внутренней среды со знаком плюс или минус, придают поступающей информации эмоционально положительную или эмоционально отрицательную субъективную окраску. В последние годы в связи, главным образом, с изучением механизмов эмоциональной памяти [21] установлена особая роль в формировании эмоций ряда ядер среднего и продолговатого мозга. Выяснилось, что в этих отделах головного мозга имеются структуры, связанные с отрицательно-эмоциональными состояниями, и структуры, связанные с положительно-эмоциональными состояниями.
К таким структурам, прежде всего, следует отнести парные ядра, расположенные в продолговатом мозгу у дна четвёртого желудочка (напомним, что этот желудочек связан сильвиевым водопроводом с третьим желудочком, находящимся в непосредственном контакте с системой гипоталамус–гипофиз) – церулярные ядра. Название происходит от латинского обозначения участков мозга, где расположены эти ядра, – locus coeruleus («голубоватое место»; в литературе часто встречается также другой термин – «синее пятно»).
Отростки нейронов, образующих церулярные ядра, отходят ко всем основным структурам головного мозга. Так, областями, где имеется большое число нервных окончаний церулярных ядер, являются кора больших полушарий, гипоталамус, таламус, гиппокамп. Сильная связь структур голубоватых мест с корой головного мозга даёт основание для того, чтобы говорить о церулярно-кортикальной системе [75]. Медиатором, обеспечивающим прохождение нервного импульса по нервным волокнам, отходящим от церулярных ядер, является норадреналин (иными словами, церулярные структуры мозга являются норадренергическими). Синтезируемый нейронами структур голубоватых мест норадреналин по отросткам аксонов, отходящих от этих нейронов, доставляется ко многим структурам мозга. Так, разрушение ядер голубоватых мест сопровождается сильным снижением уровня норадреналина в передних отделах мозга.
Норадренергические структуры выявляют обычно методом Фалька: если препарат этих структур обработать в парах формальдегида, образующие их клетки начинают флюоресцировать, они дают яркое зелёное свечение. Этот метод применяется для обнаружения и других структур с высоким содержанием биологически активных веществ группы моноаминов, к которым помимо норадреналина и адреналина, относятся дофамин, серотонин, гистамин и др. Эти вещества содержат в своей молекуле одну аминогруппу, это и определяет их название. Разные моноамины отличаются друг от друга цветом флюоресценции. В механизмах эмоций важную роль играет ещё один моноамин – серотонин, дающий при выявлении содержащих его структур методом Фалька жёлтое свечение.
Из числа ядер, синтезирующих серотонин, особенно широкими связями со структурами головного мозга обладают две группы серотонинергических нейронов, расположенных по шву (средней линии) среднего мозга: дорзальное ядро (ядро В7 в обозначениях [103, 104]) и медиальное ядро (ядро В8). Ядро В7 входит в серое вещество, окружающее сильвиев водопровод. Нервные волокна, отходящие от скопления нейронов, образующих ядра шва, оканчиваются в коре полушарий головного мозга, гипоталамусе, таламусе, гиппокампе и ряде других структур лимбической системы.
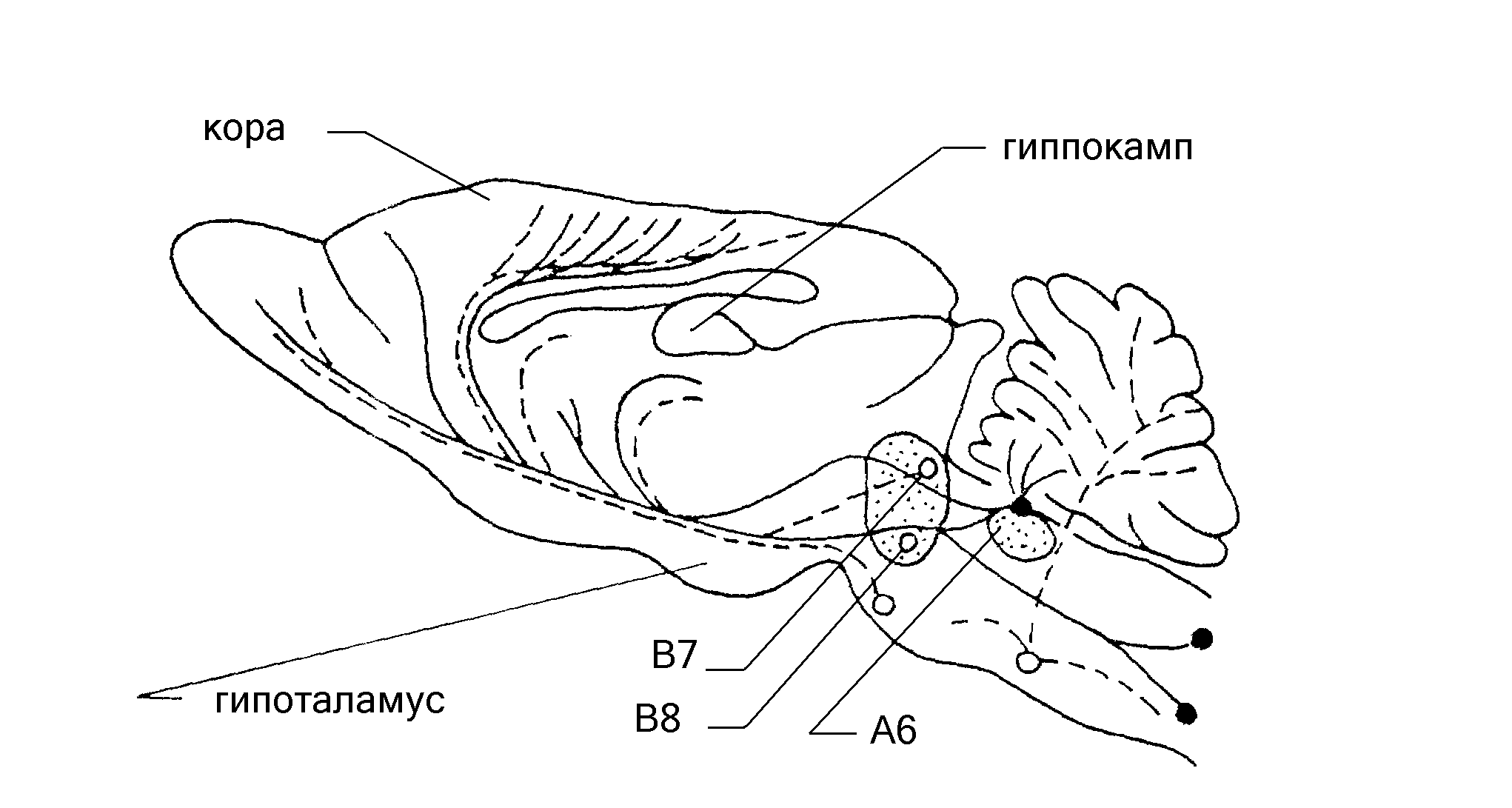
Рис.3. Схема продольного разреза головного мозга крысы с распределением серотонинергических (пунктир) и норадренергических (сплошные линии) нейронов. Кружками показаны ядра, синтезирующие серотонин, сплошными точками – ядра, синтезирующие норадреналин. Аксоны, идущие от ядра А6 (locus coeruleus) и от ядер В7 и В8 (ядра шва), достигают коры мозга и связаны с процессом обучения.
При сравнительном анализе структурной организации серотонинергической системы, связанной с ядрами шва, и норадренергической церулярной системы обнаруживается большое сходство в распределении окончаний аксонов, отходящих от обеих структур (рис.3) [21, 76, 103, 104]. Это является анатомическим указанием на то, что обе структуры оказывают сопряженное влияние на иннервируемые ими образования центральной нервной системы. Имеются и другие свидетельства сильного взаимодействия обеих структур, прежде всего – прямая взаимная иннервация: норадренергические волокна от ядер голубоватого места идут к дорзальному ядру шва (ядро В7), и, наоборот, от ядра В7 отходят аксоны, направляющиеся к синему пятну [76, 103, 104]. На функциональное взаимодействие серотонин- и норадренергических структур указывают и наличие общих ферментов, участвующих в обмене как серотонина, так и норадреналина [21].
Систематические комплексные исследования закономерностей и механизмов формирования эмоциональной памяти, выполненные под руководством Е.А.Громовой в Институте биологической физики ЦБИ АН СССР в Пущино-на-Оке в 70-80-х годах, показали, что серотонинергические структуры участвуют в процессе обучения при эмоционально положительном подкреплении, тогда как норадренергические структуры способствуют выработке реакций при обучении на основе эмоционально отрицательного подкрепления. Эти результаты (а также ряд других [21]) позволяют сделать вывод, что формирование эмоционально положительного состояния определяется преимущественно повышением активности серотонинергических систем головного мозга, а эмоционально отрицательные состояния связаны с преобладанием активности норадренергических систем [57].
Экспериментальное установление самостоятельной структурной основы положительных эмоций имеет большое концептуальное значение. До этого было широко распространено представление о положительных эмоциях как о результате удовлетворения потребности, «редукции влечения» [43, 61]. Сущность этой теории поведения целостного организма состоит в том, что возникшее у организма возбуждение при встрече с биологически значимыми факторами побуждает его к действиям для удовлетворения потребности; эти действия в случае их успешности ведут к «редукции влечения», к состоянию, воспринимаемому как «награда». С позиций теории «редукции влечения» любое состояние возбуждения рассматривали как «штраф» для нервной системы, поэтому предполагалось, что состояние удовлетворения потребности должно протекать на фоне торможения, уменьшения активности, покоя, усилия. Поэтому в экспериментах на животных о наличии положительно-эмоционального состояния долгое время судили лишь по поведенческим признакам, проявляющимся после удовлетворения какой-то потребности. Расслабленная поза животного, замедленные действия, лёгкая дремота считались признаками положительного эмоционального состояния.
Имеются однако многочисленные факты, свидетельствующие о том, что связывать положительные эмоции только с усилением тормозных процессов нельзя. Состояние повышенного настроения у человека – преимущественно активационное состояние [66]. Об этом, в частности, свидетельствуют данные В.М. Смирнова [79], который показал, что искусственно вызванным активационным состояниям у людей с вживлёнными в мозг электродами, сопровождающимся подъёмом настроения, положительными эмоциональными переживаниями, сопутствует повышение двигательной и психической активности. Положительная эмоция радости часто сопровождается двигательным возбуждением и весьма далека от состояния, при котором преобладает торможение.
Все эти и многие другие факторы (вспомним хотя бы поведение собаки, встречающей появление отсутствовавшего хозяина) находятся в противоречии с концепцией «редукции влечения»; однако они становятся естественными с точки зрения представления о самостоятельных эмоциогенных структурах, обеспечивающих положительно-эмоциональную активацию мозга.
В зависимости от значимости раздражителя в обеспечении необходимого уровня реактивности коры головного мозга может участвовать более или менее широкий круг эмоциогенных (с точки зрения активационной теории эмоций) структур. Если значимость и новизна проблемной ситуации не слишком велика, активация коры и сопряжённых с ней отделов центральной нервной системы может быть обеспечена внутрицентральными механизмами, обусловленными активностью церулярно-кортикальной системы, структур головного мозга, связанных с ядрами шва, ретикулярной формацией, гипоталамусом.
При высокой значимости раздражителя или большой длительности его воздействия интенсивное и продолжительное функционирование норадренергических ядер продолговатого и среднего мозга, гипоталамуса и ретикулярной формации ствола мозга стимулирует активность симпато-адреналовой системы. Последняя включает в себя, помимо вегетативной симпатической нервной системы, ещё и мозговое вещество надпочечников, которое в ответ на симпатическую нервную активность секретирует в кровь адреналин и норадреналин. Отметим здесь единую природу проводимости нервных импульсов в структурах, связанными с отрицательными эмоциями, и в вегетативных симпатических структурах. При высокой значимости раздражителя все норадренергические структуры центральной и периферийной нервной системы включаются одновременно (под действием одних и тех же факторов), вследствие участия церулярных ядер эта активация имеет отрицательно-эмоциональную окраску.
Вовлечение в реакцию на внешнее воздействие симпатической нервной системы приводит, как мы видели выше, к экстренной и широкой перестройке организма (усиление сердечной деятельности, повышение тонуса скелетной мускулатуры и так далее), направленной на реализацию немедленной ответной реакции.
Мышечная напряжённость, связанная с активностью симпатической нервной системы, выполняет двоякую функцию. С одной стороны, реакция на внешний раздражитель может включать мускульную активность; отсюда – усиление метаболизма в мышечных тканях, как составная часть первичной реакции организма на любую проблемную ситуацию. С другой стороны, мускульная напряжённость приводит к широкому возбуждению коры полушарий головного мозга. Импульсы, посылаемые от мышечно-нейрональных синапсов, достигают многочисленных нейронов коры головного мозга, оказывая широкие активирующие воздействия прежде всего на её соматосенсорные моторные и ассоциативные области.
Способность мускульного напряжения стимулировать нейроны головного мозга лежит, в частности, в основе воинской команды «Смирно!»: напряжение скелетной мускулатуры, когда военнослужащий принимает и поддерживает соответствующую стойку, позволяет «включить» в нужный момент структуры головного мозга, обеспечивающие точное и сознательное усвоение информации, содержащейся в последующих командах. Большую роль в оперативном управлении уровнем интенсивности мыслительной деятельности человека играет тонус мышц лица. Хорошо известны мимические реакции, связанные с мышлением, такие как нахмуренный лоб, складка между бровями, сжатые губы, играющие желваки, прищуренный взгляд. Наиболее тонкая регулировка уровня мобилизации коры головного мозга для решения проблемы, которая в данный момент занимает человека, осуществляется мышцами век: частотой моргания, прищуром. В случае, если мыслительная деятельность осуществляется на пределе возможностей мозга, моргание может становится неуправляемым, принимать форму тика.
Снижение активности обширных областей коры полушарий головного мозга в результате расслабления мускулатуры лежит в основе многих приёмов аутотренинга.
Таким образом, с позиций активационной теории эмоций имеются веские основания для того, чтобы отнести скелетную мускулатуру к эмоциогенным структурам, причём ввиду её тесной связи с норадренергической компонентой нервной системы – к структурам отрицательно-эмоциогенным.
Активация симпато-адреналовой системы, помимо возбуждения симпатической нервной системы, включает секреторную деятельность мозгового вещества, надпочечников. Этот внутренний слой надпочечников выделяет в кровь адреналин и норадреналин, которые, поступая во внутреннюю среду организма, обеспечивают те же реакции, что и симпатическая нервная система, однако в течение более длительного времени. Кроме того, адреналин, воздействуя на задние ядра гипоталамуса, участвует в запуске адаптационной реакции стресс, которая включает дополнительные механизмы мобилизации ресурсов организма для ответа на сильные и длительные воздействия [14, 49].
Заметим неравноценность положительно- и отрицательно-эмоциогенных систем. Положительные эмоции представлены преимущественно внутрицентральными механизмами воздействия на уровень мыслительной деятельности, включающими активность серотонинергических структур ядер шва и связанных с ними структур гипоталамуса и ретикулярной формации. Сила же ответа на эмоционально-отрицательные воздействия может варьировать в широких пределах от активности внутрицентральной церулярно-кортикальной системы до вовлечения в реакцию на внешнее воздействие симпато-адреналовой системы, системы гипоталамус-гипофиз, а через последнюю всей эндокринной системы организма. Такая неравноценность структурной основы положительных и отрицательных эмоций биологически целесообразна, поскольку спектр эмоционально-отрицательных воздействий чрезвычайно широк: от событий, соответствующих порогу значимости, до ситуаций, опасных для жизни. П.В. Симонов [80], исходя из предложенной им информационной теории эмоций, объясняет различие нейрофизиологических сдвигов, связанных с положительными и отрицательными эмоциями, неравноценностью мотивов при эмоциях разного знака и ограниченностью возможного прироста информации при положительных эмоциях, где естественный предел вероятности достижения цели равен единице.