

Керування руховою функцією організму
Аналіз й оцінка в нервовій системі тієї інформації, яку організм одержує через свої сенсорні системи, дозволяє йому виконувати певну діяльність. Будучи закономірною відповіддю останнього на зовнішній вплив, що здійснюється через посередництво нервової системи, вона позначається як рефлекторна реакція. Основним компонентом такої реакції є скорочення м‘язів, за допомогою чого організм може наблизитися до подразника, якщо він становить для нього інтерес, або віддалитися від подразнення, який є небезпечним, і т. ін.
Розглянемо біофізичні характеристики системної рухової діяльності вищих організмів, що лежать в основі їхніх найрізноманітніших рефлекторних реакцій.
14.1. Скелетні м‘язи як системи саморегуляції
Кожен м‘яз, якщо його розглядати відокремлено від нервових центрів, що його іннервують, поводить себе як пасивна еластична структура, здатна до пасивного розтягування під впливом зовнішньої сили. Але коли м‘яз включений у систему аферентних та еферентних зв’язків із цими центрами, його біофізичні характеристики виявляються істотно зміненими, і він набуває властивостей системи, що саморегулюється й змінює свій стан залежно від ступеня розтягнення.
Ця здатність забезпечується за рахунок так званих власних рефлексів м‘язів, рецепторні структури яких розміщені в самих м‘язах і які є сенсорами (датчиками) фізичних характеристик їхнього стану (довжини та напруги). Наявність таких механізмів дозволяє м‘язам за будь-якого положення чи руху кінцівок і тулуба зберігати певний вихідний фізичний стан, який і забезпечує надалі здійснення тих чи інших рухових реакцій.
Рецепторний апарат м‘язів. Сенсорами стану м‘яза є два типи м‘язових рецепторів: м‘язові веретена та сухожилкові органи. М‘язові веретена – це більш тонкі та короткі м‘язові волокна, що зібрані у веретеноподібні пучки та вкриті зовні сполучнотканинною капсулою. Головною особливість таких інтрафузальних м‘язових волокон є те, що вони поряд з руховою інервацією, яка здійснюється особливими тонкими руховими нервовими волокнами (що позначаються як гамма-волокна, оскільки вони, за класифікацією Дж. Ерлангера та Г. Гассера, за своїм діаметром належать до волокон типу А) несуть на своїй поверхні також механочутливі аферентні волокна. Кінцеві розгалуження останніх обвивають у вигляді спіралі інтрафузальні м‘язові волокна. За своєю тонкою структурою ці рецептори поділяються на два типи. Одні з них утворені аферентними волокнами більшого діаметра (так званої групи Iа) і обплітають центральну частину м‘язового волокна (первинні або ануло-спіральні закінчення), другі – аферентними волокнами меншого діаметра (група II), що розташовані на інтрафузальних волокнах більш периферійно (вторинні закінчення).
Мембранні механізми активації цих сенсорних структур, як і інших механорецепторів, ще не з‘ясовано, мабуть, вони пов‘язані з розтягненням їхньої мембрани при розтягненні інтрафузальних волокон, на яких вони розташовані, й активації при цьому механочутливих мембранних іонних каналів, струм через які викликає мембранну деполяризацію. Проте характер перетворення сигналу, що при цьому відбувається, вивчено дуже детально завдяки відведенню імпульсації від відповідних аферентних волокон. Приклади такої імпульсації наведено на рис. 14.1, б. У первинних закінченнях розтягнення м‘яза викликає розряд імпульсів, частота якого залежить як від ступеня розтягнення, так і від його швидкості. Такі рецептори функціонують як пропорційно-диференційні датчики довжини м‘яза. У вторинних закінченнях, що є значно менш чутливими до розтягнення, імпульсний розряд відбиває лише ступінь останнього (пропорційні датчики довжини м‘яза). Характеристики цих датчиків добре виявляються в тому разі, якщо м‘яз не розтягується пасивно, а активно
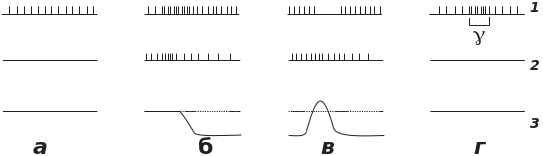
Рис. 14.1. Приклади імпульсної активності аферентів рецепторів м’язових веретен (1) та сухожилкових рецепторів (2) у спокої (а), при розтягуванні м’яза (в), при його ізотонічному скороченні (б) та при скороченні інтрафузальних м’язових волокон активацією мотонейронів (г). Рядок 3 ілюструє зміни напруження мяза
скорочується під впливом нервового сигналу, що надходить з рухового нервового волокна. При цьому імпульсація від м‘язового веретена, якщо її було викликано попереднім розтягненням, припиняється (рис. 14.1, в), є очевидним наслідком тієї обставини, що інтрафузальні волокна розташовані паралельно звичайним м‘язовим волокнам, і тому при скороченні м‘яза їхня довжина зменшується.
Істотні зміни аферентної сигналізації спостерігаються тоді, коли до м‘яза надходять відцентрові імпульси по гамма-волокнах, що іннервують інтрафузальні м‘язові волокна. Оскільки кількість останніх у загальній масі м‘язової тканими є незначною, то їхнє скорочення не може викликати зміни напруги м‘яза. Але підвищується чутливість розташованого на них рецепторного апарату до розтягнення, і відповідно за одного й того самого його ступеня частота виникаючих аферентних імпульсів стає вищою, ніж це має місце за відсутності впливу імпульсації з гамма-волокон (рис. 14.1, г). Причиною такого ефекту є саме скорочення інтрафузальних волокон і відповідне зменшення їхньої довжини. Тому відповідно розтягнення всього м‘яза більш ефективно збуджуватиме розміщений на цих волокнах рецепторний апарат.
Таким чином, у рецепторах м‘язових волокон закладено механізм регуляції їхньої чутливості, який керується сигналами із центральної системи. Така система може компенсувати ті зміни в чутливості м‘язового рецепторного апарату, які мають виникати при зміні фізичного стану м‘яза – його розтягненні чи вкороченні залежно від положення місць його прикріплення до кісткової системи, а також розтягненні чи вкороченні інтрафузальних м‘язових волокон. Точність такого регулювання посилюється тією обставиною, що сигнали з боку гамма-волокон можуть бути двох типів – динамічними та статичними. Перші змінюють чутливість м’язових рецепторів до швидкості розтягнення м’яза, разом з тим мало впливаючи на їхню стаціонарну чутливість; другі ж – ефективно змінюють частоту аферентних сигналів саме за постійного розтягненні м‘яза. Таким чином, регулюютьсяяк пропорційні, так і диференційні характеристики функціонування м’язових рецепторів.
Сухожилкові рецептори Гольджі, як на це вказує їхня назва, розташовані серед сполучнотканинних волокон сухожилків, за допомогою яких м‘язи прикріплюються до кісток. Вони є терміналями відповідних аферентних волокон, що належать до групи Iв. Їхні характеристики як датчиків зовнішніх подразнень може бути чітко продемонстровано, як і в попередньому випадку, при реєстрації тієї аферентної імпульсації, яку вони генерують у відповідних волокнах при розтягненні м‘яза чи його скороченні. При розтягненні м‘яза частота цієї імпульсації зростає, як це має місце у пропорційних рецепторів м‘язових веретен, але й при скороченні м‘яза імпульсація не тільки не зникає, але й посилюється пропорційно силі скорочення (рис. 14.1, в). Це вказує на те, що відповідні рецептори розташовані не паралельно, а послідовно з м‘язовими волокнами й служать індикаторами напруження м‘яза, а не його довжини, як це характерно для рецепторів м‘язових веретен.
Механізми саморегуляції стану м‘яза. Описаний вище складний рецепторний апарат є основою саморегуляторного механізму, надзвичайно важливого для успішного функціонування скелетної мускулатури. Цей механізм включає, поряд з описаною вище системою рецепторів й аферентних волокон, відповідні нейронні структури спинного мозку, до яких надходять аферентні сигнали і в яких вони переходять на рухові (моторні) нейрони, що надсилають еферентні (рухові) команди до тих самих м‘язів. Створена таким перемиканням рефлекторна дуга відзначається простотою й включає в себе послідовне з‘єднання всього двох нервових клітин – аферентної (чутливої), сома якої розташована в задньокорінцевому ганглії, та еферентної (рухової), розташованої в передньому розі спинного мозку. Відповідно вона позначається як двонейронна (моносинаптична).
Коли при розтягненні м‘яза відбувається активація рецепторів м‘язових веретен, це приводить до виникнення через зазначену моносинаптичну рефлекторну дугу до збудження рухових нейронів цього м‘яза та інервованих ними нервово-м‘язових одиниць (рис. 14.2). М‘яз скорочується, тим самим протидіючи його розтягненню. Навпаки, при скороченні м‘яза відбувається зменшення чи повне виключення аферентної імпульсації від цих рецепторів; синаптична дія на рухові нейрони послаблюється й відповідно зменшується сила скорочення м‘яза.
Таким чином, за принципом сервомеханізму, відбувається автоматична компенсація змін навантаження на м‘яз, спрямована на збереження його постійної довжини (розтягнення). Аферентні сигнали від первинних рецепторів м‘язових веретен відіграють при цьому основну роль, оскільки вони мають гостро спрямований характер і впливають, головним чином, на активність рухових нейронів того самого (гомонімного) м‘яза. Рефлекторні впливи від вторинних рецепторів виявляються більш дифузними і захоплюють рухові нейрони кількох м‘язів. Функціональний зміст цих впливів є менш чітким.

Рис. 14.2. Схема взаємодії низхідної рухової команди з власним рефлекторним механізмом м’яза:
1 – низхідні рухові шляхи спинного мозку; 2 – мотонейрони; 3 – екстра- та інтрафузальні м’язові волокна
Аферентні волокна від сухожилкових рецепторів не встановлюють прямих (моносинаптичних) контактів з руховими нейронами, а справляють власний вплив через посередництво короткоаксонних проміжних нейронів спинного мозку. Характер цих впливів є протилежним тим, які справляють описані вище сигнали від рецепторів м‘язових веретен: у мотонейронах гомонімних м‘язів виникають уже згадані гальмівні постсинаптичні потенціали (ГПСП), які пригнічують їхню збуджувальну дію. Оскільки такого гальмування зазнає той самий м‘яз, який розтягується, останнє позначається як аутогенне гальмування.
Оскільки сухожилкові рецептори, як це зазначалося вище, є датчиками напруження м‘яза, то зрозуміло, що їхня активація спрямовуватиметься на зниження цього напруження при м‘язовому скороченні. І, навпаки, при зниженні м‘язового тонусу відбуваються розгальмовування відповідних рухових нейронів і підвищення їхньої активності. Таким чином, також створюється ще один сервомеханізм, спрямований на компенсацію змін напруження м‘яза та збереження його оптимального рівня.
Зрозуміло, що при природній скоротливій діяльності м‘язів неможливо зберегти постійними як його довжину, так і напруження. За такої діяльності обидва сервомеханізми виявляються, по суті, протидіючими. Вірогідно, що при цьому на постійному рівні підтримується, швидше, не довжина чи напруження м‘яза, а його більш інтегральна характеристика, що може бути позначена як жорсткість (англ. stiffness), тобто відношення зміни напруження м‘яза до зміни його довжини. Зазвичай таке відношення досягає 75 (тобто для подовження м‘яза вдвічі необхідно прикласти силу, що в 75 разів перевищує силу, яку м‘яз розвиває при його тетанічному скороченні).
Жорсткість м‘язів має велике значення, оскільки вона визначає їхню поведінку за невеликих змін їхньої довжини, що зазвичай має місце в умовах фізіологічної діяльності. Так, при підтриманні пози тіла вона забезпечує стабілізацію взаємного положення окремих частин тіла та кінцівок за різних відхиляючих впливів. Таке регулювання стану м‘язів позначається як підтримання їхньої короткодіапазонної жорсткості.