

7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 325
Рис. 7.2. Схема синтеза гемоглобина у человека в эритроидной клетке.
Указаны ферменты, контролирующие этапы синтеза молекулы гема в митохондриях, а также хромосомы, ответственные за синтез цепей глобина А (β-цепи) и гемоглобинов плода (γ- и δ-цепи). Пояснения в тексте.
Соединение гемоглобина с молекулой О2 называется оксигемоглобином. Сродство гемоглобина к кислороду и диссоциация оксигемоглобина (отсо-единения молекул кислорода от оксигемоглобина) зависят от напряжения кислорода (РО2), углекислого газа (РСО2) в крови, рН крови, ее температу-ры и концентрации 2,3-ДФГ в эритроцитах. Так, сродство повышают уве-личение РО2 или снижение РСО2 в крови, нарушение образования 2,3-ДФГ в эритроцитах. Напротив, повышение концентрации 2,3-ДФГ, сни-жение РО2 крови, сдвиг рН в кислую сторону, повышение РСО2 и темпе-ратуры крови — уменьшают сродство гемоглобина к кислороду, тем самым облегчая ее отдачу тканям. 2,3-ДФГ связывается с β-цепями гемоглобина, облегчая отсоединение О2 от молекулы гемоглобина. Увеличение концен-трации 2,3-ДФГ наблюдается у людей, тренированных к длительной физи-ческой работе, адаптированных к длительному пребыванию в горах. Окси-гемоглобин, отдавший кислород, называется восстановленным, или дезок-сигемоглобином. В состоянии физиологического покоя у человека гемогло-бин в артериальной крови на 97 % насыщен кислородом, в венозной — на 70 %. Чем выраженней потребление кислорода тканями, тем ниже насы-
326 • Функции систем жизнеобеспечения организма
щ ение венозной крови кислородом. Например, при интенсивной физической работе потребление кислорода мышечной тканью увеличивается в несколько десятков раз и насыщение кислородом оттекающей от мышц венозной крови снижается до 15 %. Содержание гемоглобина в отдельном эритроците составляет 27,5—33,2 пикограмма. Снижение этой величины свидетельствует о гипохромном (т. е. пониженном), увеличение — о гипер-хромном (т. е. повышенном) содержании гемоглобина в эритроцитах. Этот показатель имеет диагностическое значение. Например, гиперхромия эритроцитов характерна для В,2-дефицитной анемии, гипохромия — для желе-зодефицитной анемии.
7.1.3. Старение и разрушение эритроцитов в организме
Эритроциты у человека функционируют в крови максимум 120 дней, в среднем 60—90 дней. Старение эритроцитов связано с уменьшением образования в эритроците количества АТФ в ходе метаболизма глюкозы в этой клетке крови. Уменьшенное образование АТФ, ее дефицит нарушает в эритроците процессы, обеспечиваемые ее энергией, — восстановление формы эритроцитов, транспорт катионов через его мембрану и защиту компонентов эритроцитов от окисления, их мембрана теряет сиаловые кислоты. Старение эритроцитов вызывает изменения мембраны эритроцитов: из дискоцитов они превращаются в эхиноциты, т. е. эритроциты, на поверхности мембраны которых образуются многочисленные выступы, выросты (рис. 7.3).
Причиной формирования эхиноцитов помимо уменьшения воспроизводства молекул АТФ в эритроците при старении клетки является усиленное образование лизолецитина в плазме крови, повышенное содержание в ней жирных кислот. Под влиянием перечисленных факторов изменяется соотношение поверхности внешнего и внутреннего слоев мембраны эритроцита за счет увеличения поверхности внешнего слоя, что и приводит к появлению выростов на мембране.
По степени выраженности изменений мембраны и формы эритроцитов различают эхиноциты I, II, III классов и сфероэхиноциты I и II классов. При старении эритроцит последовательно проходит этапы превращения в эхиноцит III класса, теряет способность изменять и восстанавливать дис-ковидную форму, превращается в сфероэхиноцит и разрушается. Устранение дефицита глюкозы в эритроците легко возвращает эхиноциты I—II классов к форме дискоцита. Эхиноциты начинают появляться, например, в консервированной крови, сохраняемой в течение нескольких недель при 4°С, или в течение 24 ч, но при температуре 37 °С. Это связано с уменьшением образования АТФ внутри клетки, с появлением в плазме крови лизолецитина, образующегося под влиянием лецитин-холестерол-ацетилтранс-ферразы, ускоряющих старение клетки. Отмывание эхиноцитов в свежей плазме от содержащегося в ней лизолецитина или активация в них гликолиза, восстанавливающей уровень АТФ в клетке, уже через несколько минут возвращает им форму дискоцитов.
Стареющие эритроциты становятся менее эластичными, вследствие чего разрушаются внутри сосудов (внутрисосудистый гемолиз) или же становятся добычей захватывающих и разрушающих их макрофагов в селезенке, купферовских клетках печени и в костном мозге (внесосудистый или внутриклеточный гемолиз). Внутриклеточным гемолизом в сутки разрушается 80—90 % старых эритроцитов, содержащих 6—7 г гемоглобина, из которых освобождается в макрофагах до 30 мг железа. После отщепления от гемо-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 327
4,5 6 7,2 8 9,5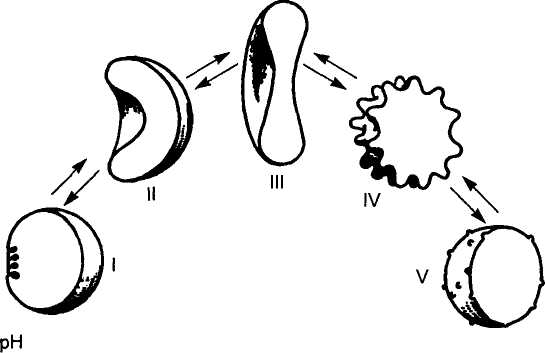
I — сферостоматоцит, II — стоматоцит, III — дискоцит, IV — эхиноцит, V — сфероэхиноцит.
глобина гем превращается в желчный пигмент билирубин, который поступает с желчью в кишечник и под влиянием микрофлоры кишечника последовательно превращается в уробилиноген, а затем в стеркобилиноген. Оба соединения выводятся из организма с калом и мочой, под влиянием света и воздуха превращаясь в стеркобилин и уробилин. При метаболизме 1 г гемоглобина образуется 33 мг билирубина.
Внутрисосудистым гемолизом разрушается 10—20 % эритроцитов. При этом гемоглобин поступает в плазму, образует с плазменным гликопротеи-ном гаптоглобином комплекс гемоглобин—гаптоглобин. В течение 10 мин 50 % комплекса поглощается из плазмы паренхиматозными клетками печени, что предупреждает поступление свободного гемоглобина в почки и тромбирование им их нефронов. У здорового человека в плазме содержится около 1 г/л плазмы гаптоглобина, что оставляет несвязанным с ним в плазме крови не более 3—10 мг гемоглобина. Молекулы гема, высвобождающиеся из связи с глобином при внутрисосудистом гемолизе, связываются белком плазмы — гемопексином, транспортируются им в печень и также поглощаются паренхиматозными клетками печени, где подвергаются ферментному разрушению до билирубина.
7.1.4. Роль ионов железа в эритропоэзе
В организме содержится 4—5 г железа в виде резервного ('/,,) и функционально-активного железа (3/4)- 62—70 % железа находится в гемоглобине эритроцитов, 5—10 % — в миоглобине, остальное — в тканях, где оно участвует во многих метаболических процессах: в составе металсодержащих энзимов—цитохромов обеспечивают митохондриальный транспорт электронов, синтез ДНК и деление клеток, метаболизм гормонов мозгового вещества надпочечников, детоксикационные механизмы, снижающие активность токсичных веществ под влиянием цитохрома Р450, в состав которого входит железо; поддерживают активность глицерол-3-фосфатдегидрогеназы в поперечно-полосатых мышцах, что сохраняет физическую работоспособность