

Физиология возбудимых тканей
26. Механизм мышечного сокращения. Мышцы состоят из мышечных волокон, которые состоят из множества тонких нитей -миофибрилл, расположенных продольно. Каждая миофибрилла состоит из протофибрилл – нитей сократительных белков актина и миозина. Перегородки, называемые Z-пластинами, разделяют миофибриллы и, следовательно, мышечное волокно на участки – саркомеры. В саркомере наблюдают правильно чередующиеся светлые и темные полосы. Эта поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера свободно расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который дает двойное лучепреломление, т. к. содержит нити миозина и актина и называется анизотропным или А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, т. к. они обладают одним лучепреломлением и называются изотропными или j-дисками. По их середине проходит темная линия - Z-мембрана. Именно благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят исчерченными (поперечно-полосатыми) (рис. 6).
В состоянии покоя концы толстых и тонких нитей лишь незначительно перекрываются на уровне А-диска. В соответствии с теорией скользящих нитей при сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера., Сами актиновые и миозиновые нити своей длины не изменяют.
Механизм скольжения нитей. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек (подобно скрученным ниткам бус) молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя нитями актина лержат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.
Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему) . Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена" система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек - терминальные цистерны –подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В этих пузырьках сосредоточено основное количество внутриклеточного кальция.
В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и глобул тропонина. При возбуждении ПД быстро распространяется по мембранам поперечной системы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина в результате чего отодвигается нить тропомиозина и открываются участки актина, присоединяющие миозиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его "сгибанию"), в результате конформации этой части миозиновои молекулы, и перемещению нити актина на один шаг( на один «гребок») к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина позволяют "грести" или тянуть актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом, из протофибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и cнoвa блoкиpyeт aктивныe центры актина. Вместе с тем происходит фосфорилирование миозина за счет АТФ, который не только заряжает системы для дальнейшей работы, но и способствует временному разобщению нитей. Удлинение (расслабление) мышцы после ее сокращения является процессом пассивным, поскольку актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон и мышцы, а также силы растяжения мышц антагонистов.
27. Гладкие мышцы. Гладкие мышцы, формирующие мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых внутренних органов, построены из веретенообразних одноядерных мышечных клеток. Отдельные клетки в гладких мышцах функционально связаны между собой низкоомными электрическими контактами – нексусами. За счет этих контактов потенциалы действия и медленные волны деполяризации беспрепятственно распространяются с одного мышечного волокна на другое. Поэтому несмотря на то, что двигатёльные нервные окончания расположены на небольшом числе мышечных волокон, вследствие беспрепятственного распространения возбуждения с одного волокна на другое в реакцию вовлекается вся мышца. Следовательно, гладкие мышцы представляют собой не морфологический, а функциональный синтиций.
Особенностью гладких мышц является их способность осуществлять относительно медленные движения и длительные тонические сокращения. Медленные, имеющие ритмический характер, сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц особенно хорошо выражены в сфинктерах полых органов, которые препятствуют выходу содержимого этих органов.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов.
Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. Эти различия хорошо наблюдать при медленном растяжении гладкой и скелетной мышцы. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Благодаря высокой пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии. Так, например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления внутри его.
Сильное и резкое растяжение гладких мышц вызывает их сокращение. Переднее обусловлено нарастающей при растяжении деполяризацией клеток, обусловливающих автоматию гладкой мышцы. Сокращение, индуцируемое растяжением, играет важную роль в авторегуляции тонуса кровеносных сосудов, а также обеспечивает непроизвольное (автоматическое) опорожнение переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах одиночное сокращение продолжается несколько секунд. Тетаническое сокращение возникает при низкой частоте слияния одиночных сокращений и низкой частоте сопровождающих его ПД.
В отличие от скелетной мышцы гладкая мышца кишки, мочеточника, желудка и матки развивает спонтанные тетанообразные сокращения в условиях ее изоляции и денервации, и даже после блокады интрамуральных ганглиев. В этом случае они возникают не в результате передачи нервных импульсов с нерва, вследствие активности клеток, обладающих автоматией, т. е. пейсмекерных клеток. Последние идентичны по структуре другим мышечным клеткам, но отличаются по электрофизиологическим свойствам. В этих клетках возникают препотенциалы или пейсмекерные потенциалы, деполяризующие мембрану до критического уровня. Вследствие входа, главным образом, ионов кальция мембрана деполяризуется до изоэлектрического уровня, а затем поляризуется с обратным знаком (до +20 мВ). Реверсия потенциала длится в течение нескольких секунд. За реполяризацией следует новый препотенциал, который вызывает еще один потенциал действия.
Вегетативная нервная система и ее медиаторы оказывают на спонтанную активность пейсмекеров модулирующие влияния. При нанесении ацетилхолина на препарат мышцы толстой кишки пейсмекерные клетки деполяризуются до околопорогового уровня и частота ПД возрастает. Инициируемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота ПД, тем сильнее суммированное сокращение. Нанесение на этот препарат норадреналина гиперполяризует мембрану и таким образом снижает частоту ПД и величину тонуса.
Возбуждение гладкомышечных клеток вызывает либо увеличение входа ионов кальция через мембрану клетки, либо высвобождение ионов кальция из внутриклеточных хранилищ. В результате повышения концентрации ионов кальция в саркоплазме активируются сократительные структуры. Так же как сердечная и скелетная мышца, гладкая мышца всегда пассивно расслабляется, если концентрация ионов кальция очень мала. Однако расслабление гладких мышц происходит более медленно, т. к. замедлено удаление ионов кальция.
28. Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда На границе возбужденного и невозбужденного участка начинает протекать электрический ток . Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения , в то время как ранее возбужденные участки возвращаются в состояние покоя. Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны . Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны . Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье . Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
• Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
• Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
29. Рецепторы играют ведущую роль в получении организмом информации о состоянии внешней и внутренней среды. Благодаря большому многообразию рецепторов человек способен воспринимать стимулы разных модальностей.
Рецепторы представляют собой конечные специализированные образования, которые предназначены для восприятия энергии раздражителя и трансформации ее в специфическую активность нервной клетки. У большинства рецепторных аппаратов основной структурной единицей является клетка, снабженная подвижными волосками или ресничками, которые представляют собой как бы периферические подвижные антенны. В составе волосков выделяют 9 пар периферических фибрилл, которые сокращаются под действием АТФ, благодаря чему осуществляются непрерывные поиски адекватного стимула и обеспечиваются условия взаимодействия с ним. Центральные 2 фибриллы выполняют азотистую функцию. Общий механизм рецепции слагается из механохимических молекулярных процессов, обеспечивающих движение антенн, и общих биохимических циклов при взаимодействии специфического стимула с рецепторными мембранами антенн. Однако, у некоторых рецепторов во взаимодействии со стимулом принимает участие вся клетка (например, хеморецепторные клетки, чувствительные к напряжению кислорода в 'крови), у других восприятие осуществляется микроворсинками (вкусовые луковицы). У большинства рецепторов кожи, внутренних органов и мышц участки преобразования стимула находятся в окончаниях нервных волокон.
КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ.
В основу классификации рецепторов положено несколько критериев.
* Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
* Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осморецепторы и др.
* Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
* Отношение к одной или нескольким модальностям: моно- и полимодальные
(мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.) .
* Способность воспринимать раздражитель, находящийся на расстоянии от
рецептора или при непосредственном контакте с ним: контактные и дистантные.
* Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
* Скорость адаптации: быстроадаптирующиеся (тактильные),
медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
* Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
* Морфофункциональная организация и механизм возикновения возбуждения:
первичночувствующие и вторичночувствующие.
В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) - потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
Механизм возникновения возбуждения у этих рецепторов различен. В первичночувствующем рецепторе трансформация энергии раздражителя и возникновение импульсной активности идет в самом сенсорном нейроне. У вторичночувствующих рецепторов между сенсорным нейроном и стимулом расположена рецептирующая клетка, в которой под влиянием раздражителя идут процессы трансформации энергии раздражителя в процесс возбуждения. Но в этой клетке не возникает импульсной активности. Рецепторные клетки синапсами соединены с сенсорными нейронами. Под влиянием потенциала рецептирующей клетки выделяется медиатор, который возбуждает нервное окончание сенсорного нейрона и вызывает в нем появление локального ответа - постсинаптического потенциала. Он оказывает деполяризующее действие на отходящее нервное волокно, в котором возникает импульсная активность.
Следовательно, у вторичночувствующих рецепторов локальная деполяризация возникает дважды: в рецептирующей клетке и в сенсорном нейроне. Поэтому принято называть градуальный электрический ответ рецептирующей клетки рецепторным потенциалом, а локальную деполяризацию сенсорного нейрона генераторным потенциалом, имея ввиду, что он генерирует в отходящем от рецептора нервном волокне распространяющееся возбуждение. У первичночувствующих рецепторов рецепторный потенциал является и генераторным. Таким образом, рецепторный акт можно изобразить в виде следующей схемы.
Для первичночувствующих рецепторов:
* I этап - специфическое взаимодействие раздражителя с мембраной рецептора;
* II этап - возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором в результате изменения проницаемости мембраны для ионов калия (или кальция);
* III этап - электротоническое распространение рецепторного потенциала к
аксону сенсорного нейрона (пассивное распространение рецепторного потенциала вдоль нервного волокна называется электротоническим);
* IV этап - генерация потенциала действия;
* V этап - проведение потенциала действия по нервному волокну в ортодромном направлении. Для вторичночувствующих рецепторов:
* I-III этапы совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране;
* IV этап - выделение медиатора пресинаптическими структурами рецептирующей клетки;
* V этап - возникновение генераторного потенциала на постсинаптической мембране нервного волокна;
* VI этап - электротоническое распространение генераторного потенциала по нервному волокну;
* VII этап - генерация потенциала действия электрогенными участками нервного волокна;
* VII этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Функциональная мобильность. Анализаторные системы способны изменять свою деятельность путем изменения количества функционирующих рецепторов в зависимости от условий окружающей среды и функционального состояния организма. Например, количество функционирующих вкусовых рецепторов больше в состоянии голода, а после приема пищи их количество уменьшается. При снижении температуры окружающей среды коичество холодовых рецепторов кожных покровов увеличивается.
Физиология ЦНС
30. Нейрон-(греч. neuron – жила, нерв) - это основная структурно-функциональная единица нервной системы, обладающая специфическими проявлениями возбудимости.
Классификация нейронов функциональная: Афферентные (чувствительные);
Вставочные (ассоциативные);
Эфферентные (двигательные);
Секреторные.
Виды нейронов:
Униполярные![]() Структура нейрона
:
Структура нейрона
:
Псевдоуниполярные
![]()
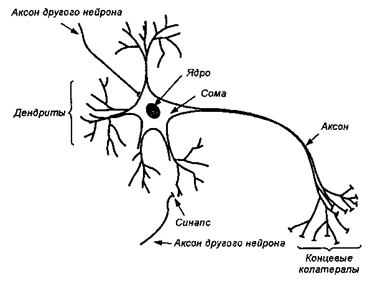
Б иполярные
иполярные![]()
Мультиполярные
Изополярные
Гетерополярные
Аксон начинается от тела клетки аксонным холмиком,по аксону передаются нервные импульсы к другим нейронам и эффекторным клеткам.Нейрон имеет большое число дендритов, на которых локализуются большинство синапсов нейрона,обеспечивающих восприятие информации от других нервных клеток.
Для возбуждения нейрона (возникновения потенциала действия) необходимы потоки афферентных импульсов и их взаимодействие. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль Са2+,причем количество медиатора прямо пропорционально входу ионов Са2+,при этом четыре иона Са2+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени по различным входам в результате пространственной суммации возбуждающего постсинаптического потенциала возникает ПД нейрона –мишени. В большинстве центральных нейронов ПД возникает в области мембраны аксонного холмика, возбудимость которой в два раза выше других участков и отсюда возбуждение распространяется по аксону и телу клетки. Такой способ возбуждения нейрона важен для осуществления его интегративной функции, т. е. способности суммировать влияния, поступающие на нейрон по разным синаптическим путям. Степень возбудимости разных участков нейрона неодинакова, она самая высокая в области аксонного холмика, в области тела нейрона она значительно ниже, и самая низкая у дендритов.
Интегративная функция нейрона (переработка информации). Структура,где начинает формироваться ПД ,- аксонный холмик.
31. Физиологические свойства нервных центров. Нервным центрам свойственно явление суммации возбуждений. Различают два вида суммации: Временная (последовательная)–обусловлена тем,что ВПСП (возбуждающий постсинаптический потенциал) от предыдущего импульса еще продолжается ,когда приходит следующий импульс. Пространственная суммация – связана с такой особенностью распространения возбуждения,как конвергенция,т. е. схождение многих афферентных путей к одному и тому же центру. Нервным центрам присуща трансформация ритма возбуждения – изменение числа импульсов ,возникающих в нейронах центра на выходе,относительно числа импульсов, поступающих на вход данного центра.Трансформация ритма возбуждения возможна в сторону как увеличения , так и уменьшения. Уменьшение числа импульсов в нервном центре объясняется снижением возбудимости последнего за счет процессов пре- и постсинаптического торможения ,а также избыточным потоком афферентных импульсов.Увелечение числа импульсов ,возникающих в центре в ответ на афферентную импульсацию ,способствуют иррадиация процесса возбуждения и последствие. Посттетаническая потенциация - явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Оно обусловленно сохранением определенного уровня ВПСП на нейроне центра, что облегчает проведение последующих возбуждений через синапсы. Нервные центры обладают быстрой утомляемостью, низкой лабильностью, высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам. Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
32. Основные принципы распространения возбуждения в ЦНС. Конвергенция - схожденеи возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу. Объясняется это наличием многих аксонных коллатералей/,вставочных нейронов,а также тем, что афферентных путей в несколько раз больше,чем эфферентных нейронов. Дивергенция (ветвление аксонов нейронов) – расширяет сферу действия каждого нейрона. В нейронных цепях,в рефлекторных дугах одностороннее распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона (но не обратоно),что объясняется свойствами химических синапсов,которые проводят возбуждение только в одном направлении. В нейронных цепях наблюдается иррадиация ,что объясняется ветвлением аксонов (дивергенция) и их способностью устанавливать многочисленные связи с другими нейронами,наличием вставочных нейронов ,аксоны которых также ветвятся. Мультипликация,реверберация??
33. Торможение в ЦНС. Торможение - активный нервный процесс ,результатом которого является прекрвщение или ослабление возбуждения. Торможение в ЦНС открыл И М Сеченов ,в опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Гольц обнаружил проявление торможения шагательного рефлекса у спинальной собаки при механическом раздражении хвоста . Наличие специальных тормозных структур в продолговатом мозге докозал Мегун,в опытах на кошках при изучении разгибательного рефлекса он установил,что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга. По отношению к синапсу торможение бывает- пресинаптическое и постсинаптическое. Пресинаптическое торможение обусловлено наличием вставочных тормозных нейронов, которые формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению, например, к мотонейрону. В любом случае активации тормозного интернейрона, он вызывает деполяризацию мембраны афферентных терминалей, ухудшающей условия проведения по ним ПД, что таким образом уменьшает количество выделяемого ими медиатора, и, следовательно, эффективность синаптической передачи возбуждения к мотонейрону, что уменьшает его активность. Постсинаптическое торможение развивается в условиях, когда медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется. Постсинаптическое торможение может быть деполяризационным, если в его основе лежит процесс длительной деполяризации, и гиперполяризационным, если - гиперполяризации.
34. Основные принципы координационной деятельности ЦНС: Координационная деятельность ЦНС – это соглосование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Принцип реципрокности (сочетанности, сопряженности, взаимоисключения). Он отражает характер отношений между центрами ответственными за осуществление противоположных функций (вдоха и выдоха, сгибание и разгибание конечности и т.д.). Например, активация проприорецепторов мышцы-сгибателя одновременно возбуждает мотонейроны мышцы-сгибателя и тормозит через вставочные тормозные нейроны мотонейроны мышцы-разгибателя. Принцип облегчения - он проявляется в том, что суммарный ответ организма при одновременном действии двух относительно слабых раздражителей будет больше суммы ответов, полученных при их раздельном действии. Причина облегчения связана с тем, что аксон афферентного нейрона в ЦНС синаптирует с группой нервных клеток, в которой выделяют центральную (пороговую) зону и периферическую (подпороговую) "кайму". Нейроны, находящиеся в центральной зоне, получают от каждого афферентного нейрона достаточное количество синаптических окончаний, чтобы сформировать потенциал действия. Принцип окклюзии - этот принцип противоположен пространственному облегчению и он заключается в том, что два афферентных входа совместно возбуждают меньшую группу мотонейронов по сравнению с эффектами при раздельной их активации, причина окклюзии состоит в том, что афферентные входы в силу - конвергенции отчасти адресуются к одним и тем же мотонейронам, которые затормаживаются при активации обоих входов одновременно . Принцип обратной связи - управление нервным центром или рабочим органом с помощью афферентных импульсов,поступающих от них. Принцип общего конечного пути -
эффекторные нейроны ЦНС (прежде всего мотонейроны спинного мозга), являясь конечными в цепочке состоящей из афферентных, промежуточных и эффекторных нейронов, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору). Например, на мотонейронах передних рогов спинного мозга, иннервирующих мускулатуру конечности, оканчиваются волокна аффрентных нейронов, нейронов пирамидного тракта и экстрапирамидной системы (ядер мозжечка, ретикулярной формации и многих других структур). Принцип доминанты- доминантное состояние двигательных центров обеспечивает автоматизированное выполнение двигательных актов ,например,в процессе трудовой деятельности человека,при выполнении гимнастических элементов.
35.Современные представления об интегральной деятельности ЦНС. Интегративная роль нервной системы заключается в объединении органов и систем в единую функциональную систему- организм. Такое объединение становится возможным благодаря трем факторам: 1) участию ЦНС в управлении опорнодвигательным аппаратом с помощью соматической нервной системы .2) регуляции функций всех тканей и внутренних органов с помощью ВНС и эндокринной системы . 3) наличию прямых и обратных (афферентных) связей ЦНС со всеми соматическими и вегетативными эффекторами. Можно выделить четыре основных уровня ЦНС: первый уровень – нейрон. Благодаря множеству возбуждающих и тормозящих синапсов на нейроне ,он превратился в ходе эволюции в решающее устройство. Второй уровень - нейрональный ансамбль (модуль) ,обладающий качественно новыми свойствами, отсутствующими у отдельных нейронов – один и тот же модуль может участвовать в деятельности различных центров и отделов ЦНС. Третий уровень – нервный центр ,нервные центры часто выступают как автономные командные устройства,реализующие управление тем или иным процессом на перефирии в составе саморегулирующейся,самовосстанвливающейся,самовоспроизводящейся системы – организм. Четвертый уровень – высший уровень интеграции,включающий все центры регуляции,объединяемые корой большого мозга в единую регулирующую систему,а отдельные органы и системы - в единую физиологическую систему – организм,подсистемы которого постоянно формируют функциональные системы.
36. Системная организация функций мозга по принципу взаимодействия проекционных, ассоциативных, интегративно-пусковых систем. Интегративные системы ствола мозга включают ретикулярную формацию, голубое пятно,ядро шва, центральное серое вещество и черную субстанцию. Ретикулярная формация представляет собой скопление нейронов,различных по функции и размерам,связанных множеством нервных волокон ,проходящих в разных направлениях и образующих сеть на всем протяжении ствола мозга,что определяет ее название. Функции черной субстанции:участвует в регуляции мышечного тонуса, координации жевания и глотания,мелких движений конечностей,в осуществлении эмоций. Функции голубого пятна: расположено в среднем мозге,нейроны гп являются адренергическими,на их мембране имеются рецепторы к адреналину и норадреналину,серотонину,ГАМК,глицину,активация которых вызывает преимущественно торможение,а также рецепторы к ацетилхолину,активация которых вызывает возбуждение. Иннервируя ядра гипоталамуса,нейроны Гп участвуют в развитии общего адаптационного синдрома при действии на организм стрессоров. Норадренергическая система контролирует болевую и неболевую импульсации сенсорных и двигательных систем,угнетая фоновую активность нейронов и их ядер,Гп участвует в регуляции деятельности висцеральных систем ,эмоциональных сосотояний(тоска,страх) ,механизмов памяти и внимания,индуцирует фазу быстрого сна. Серотонинергическая система включает в себя центр-е серое вещество ядро шва,нейроны этой системы наиболее активны во время бодрствования,эта система тормозит нейроны сенсорных,особенно болевых,путей спинного мозга,ствола и таламуса. Серотонические нейроны тормозят также симп-ие и парасим-ие нейроны спинного мозга,но оказывают возбуждающее действие на его мотонейроны,тормозят сексуальное и агрессивное поведение,потребление воды и пищи. Ассоциативная, проекционная система-???? Функциональный элемент мозга - нервная клетка.
37. Роль спинного мозга в процессах регуляции деятельности опроно-двигательного аппарата и вегетативных функций организма. Соматические рефлексы сп мозга- в основном сгибательные и разгибательные рефлексы.Защитные рефлексы возникают при раздражении кожи и тела. Рефлексы конечностей : Сгибательные – бывают фазные – однократоное сгибание конечности при однократном раздражении кожи или мышечных рецепторов; и тонические – учсатвуют в в поддержании позы. Разгибательные – моносинаптические ,возникают с проприорецепторов мышц –разгибателей: фазные возникают в ответ на однократное раздражение только мышечных рецепторов; тонические –длительное сокращение мышц разгибателей при длительном их растяжении и возбуждении мышечных рецепторов. Ритмические рефлексы- выражаются в многократном повторном сгибании конечностей. Рефлексы позы спинального рганизма - заключаются в в перераспределении тонуса мышц конечностей при изменении положения шеи относительно туловища. С помощью проводниковой функции сп мозга вышележащие отделы ЦНС получают информацию от тела организма и внутренних органов ,регулируют функции внутренних органов,эта функция осуществляется с помощью восходящих и нисходящих путей белого вещества. Спинальное животное – это животное у которого спинной мозг отделен от головного мозга, перерезка спинного мозга производится ниже 3-го шейного позвонка. Перерезка выше 3-го шейного позвонка несовместима с жизнью, потому что на уровне 1-2 шейных позвонков лежат нервные центры дыхательной мускулатуры и, если их разрушить, животное погибнет от паралича дыхательных мышц, т.е. асфиксии. Клинически важные спинальные рефлексы: локтевой – проприоцептивный,выражается в сгибании руки в локтевом суставе,возникает при ударе молоточком по сухожилию;,пяточный рефлекс - проприоцептивный, выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы мышцы голени ,возникает при ударе по ахиллову сухожилию;
38. Спинальные механизмы регуляции мышечного тонуса и фазных движений. Спинальный шок: в эксперементе тонус мышц исследуют после перерезки сп мозга,сразу после перерзки или повркждения сп мозга наблюдаются мышечная атония и отсутствие рефлексов .Главной причиной спинального шока является выключение влияния вышележащих отделов ЦНС на спинной мозг. Гипертонус мышц:Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения сп мозга усиливаются. Афферентная импульсация от мышечных рецепторов по задним корешкам сп мозга ,которая ведет к возбуждению α- мотонейронов сп мозга и сокращению скелетных мышц и развитию гипертонуса,поддерживается двумя способами 1) спонтанной активностью мышечных рецептров 2) спонтанной активностью γ- мотонейронов.
39. Продолговатый мозг и и мост. Продолговатый мозг – является продолжением спинного мозга,по структуре он сложнее,чем спинной; за счет своих ядерных образований и ретикулярной формации,участвует в реализации вегетативных ,соматических ,слуховых ,вкусовых ,вестибулярных рефлексов. Особенностью является то,что его ядра возбуждаясь обеспечивают выполнение сложных рефлексов,требующих последовательного включения разных мышечных групп. В продолговатом мозге расположены ядра следующих черепных нервов: 1) преддверноулитковый 2) языкоглоточный 3) блуждающий 4) добавочный 5) подъязычный ; Функции: 1) проводниковая - через пр. мозг проходят все восходящие и нисходящие пути спинного мозга,заканчиваются пути в коре большого мозга. 2) рефлекторные делятся на: - жизненно-важные: дыхательные и сосудистодвигательные центры ; - нежизненно-важные: защитные рефлексы (рвота,чихание), пищевого поведения,рефлексы поддержания позы. 3) Сенсорная регулирует: - рецепцию кожной чувствительности лица,в сенсорном ядре тройничного нерва; - первичный анализ рецепции вкуса: в ядре языкоглоточного нерва; - рецепцию слуховых раздражений :в ядре улиткового нерва; -рецепцию вестибулярных раздражений : в верхнем вестибулярном ядре. Мост - располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции. В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва). Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
41. Децеребрационная ригидность- повышение тонуса всех мышц, чаще с резким преобладанием тонуса мышц-разгибателей в результате нарушения связей и разобщения головного мозга и мозгового ствола на уровне среднего мозга. (при отделении красного ядра среднего мозга от нижележащих структур повышается тонус экстензоров которые контролируются вестибулярными ядрами продолговатого мозга. )
Установлено что нейроны красного ядра вызывают в основном возбуждение мышц-сгибателей. Поэтому при его нарушении преобладают влияния вестибулярных ядер и возникает децеребрационная ригидность.
В среднем мозге находятся красные ядра которые вызывают активацию альфа-мотонейронов сгибателей и тормозят активацию альфа-мотонейронов разгибателей спиного мозга
В продолговатом мозге находятся вестибулярные ядра которые активацию альфа-мотонейронов разгибателей и тормозят активацию альфа-мотонейронов сгибателей спиного мозга.
Благодаря этому красные ядра вместе с вестибулярными ядрами участвуют в регуляции позы.
42. ствол мозга обеспечивает подержание равновесия в условиях покоя и ускорения.
Это проявляется в наличие статической и стато-кинетических рефлексов. Впервые их подробно исследовал голландский физиолог Р.Магнус (рефлексы Магнуса).
Статические рефлексы условно делят на позно-тонические и выпрямительные. Оба вида рефлексов возникают в результате раздражения рецепторов вестибулярного аппарата, проприорецепторов мышц и рецепторов фасции шеи, а так же (выпрямительных) при активации рецепторов кожи. структура участвующая реализации этих рефлексов вестибулярные ядра. красные ядро и ретикулярная формация также причастна к этим рефлексам. Позно-тонические рефлексы возникаю при одновременном возбуждении рецепторов шейных мышц и вестибулярного аппарата. У животных и человека эти рефлексы хорошо выявляются в раннем возрасте. У грудных детей наблюдается лабиринтный тонический рефлекс: у ребенка лежащего на спине, повышен тонус разгибателей шеи, спины , ног если же ребенка перевернуть на живот , то увеличится тонус сгибателей шеи, спины , конечностей. Тормозится примерно к 2-3 месяцам.
Эти рефлексы у взрослых выявить трудно но при выполнении быстрых и резких движении можно выявить эти рефлексы.
Выпрямительный рефлекс проявляется в том что животное из непривычного , несвойственного положения переходит в естественное положение. (феномен кошки на 4 лапах)
Стато-кинетические рефлексы направлены на сохранения позы и ориентации в пространстве при изменении скорости движения. К стато-кинетическим рефлексам относят лифтные рефлексы: при подъеме вверх повышается тонус сгибателей ног, а при опускании вниз возрастает тонус разгибателей.
Среди рефлексов ствола мозга необходимо отметить сторожевой и ориентировочный рефлексы. Они осуществляются с участием нейронов тектума: в ответ на зрительный сигнал или на звуковой происходит активация соответствующих рецепторов, сигнал достигает первичных центров (зрительный – передние или верхние бугры четверохолмия, слуховые – задние или нижние бугры),переключение на тектоспинальные тракт – возникают ориентировочные рефлексы: поворот головы в ту сторону, откуда поступает сигнал, и одновременно происходит перераспределение тонуса мышцы, в том числе за счет изменения положения головы, благодаря которому сохраняются равновесие и возникают благоприятные условия для совершения нападения на врага или убегания.
43. У мозжечка имеется кора. Она содержит 3 слоя – молекулярный, ганглиозный и гранулярный. Белое вещество мозжечка содержит ядро шатра, пробковидное и шаровидное ядра, зубчатое ядро. Принцип работы мозжечка заключается в следующем: поступает обширная информация, в том числе от вестибулярного аппарата, от мышечных рецепторов, от кожных рецепторов, а также от коры больших полушарий. Эта информациям обрабатывается в коре мозжечка. Результаты обработки подаются на ядра мозжечка, которые управляют деятельностью красного ядра, вестибулярного ядра, ретикулярной формации. Кроме того, информация идет в кору больших полушарий, где используется для составления точных программ выполнения сложных движений. Кора мозжечка представлена 6 типами клеток: 1 клетки-зерна они являются возбуждающими в отличие от остальных 5 видов клеток, которые выполняют роль тормозных структур. 2 клетки гольджи. 3 клетки Пуркинье или грушевидные клетки самые большие клетки мозжечка, тормозные их аксон единственный выход из коры мозжечка на его ядра.4 корзинчатые клетки в молекулярном слое. 5 звездчатые клетки клетки в молекулярном слое. 6 клетки Лугаро клетки в молекулярном слое. В мозжечок поступает 2 типа волокон- лиановидные или лазящие и моховидные.
С точки зрения функций которые выполняют мозжечок делят на 3 части: архиоцеребеллум , палеоцеребеллум, неоцеребеллум.
Кора архиоцеребеллума связана с ядром шатра. Это ядро регулирует активность вестибулярных ядер. Является вестибулярным регулятором: изменяя активность нейронов вестибулярных ядер, мозжечок тем самым влияет на процессы равновесия и все, что связано с деятельностью вестибулярных ядер. Ядро шатра влияет и на нейроны ретикулярной формации моста.
Палеоцеребеллума это взаимная координация позы и целенаправленного движения а также коррекция выполнения сравнительно медленных движений на основе механизма обратной связи. Эта функция реализуется с участия двух ядер мозжечка – пробковидного и шаровидного. они влияют на деятельность красного ядра и ретикулярной формации, расположенной в продолговатом мозге. Эта часть мозжечка работает на основании информации от мышечных рецепторов и двигательной коры. Функция коррекции медленных движений имеет важное значение в процессе обучения но она не может использоваться при выполнении быстрых и сложных движений .
Неоцеребеллум - играет роль в программировании сложные движений, выполнение которых идет без использования механизма обратной связей. Информация в неоцеребеллум поступает от ассоциативных зон коры, которая вначале доставляется в нейрон моста, откуда по понтоцеребеллярному пути она поступает в неоцеребеллум. От нейронов коры можечка информация идет на зубчатое ядро. От этого ядра информация идет через таламус к двигательной коре, откуда она поступает по пирамидному и экстрапирамидному пути к альфа-мотонейронам спинного мозга. В итоге возникает целенаправленное движение, выполняемое с большой скоростью (игра на фортепьяно)
Ядра мозжечка находятся под влияниям нейронов Пуркинье. Когда активность этих тормозных нейронов возрастает, влияние ядер мозжечка на стволовые структуры уменьшается. Когда активность нейронов Пуркинье снижается, снимается их тормозное действие на ядра мозжечка и тем сам мозжечок более активно влияет на функции стволовых структур.
Мозжечок имеет важное значение в регуляции вегетативных функций организма. Мозжечок играет как и симпатическая нервная система, адаптационно-трофическую роль, т. е. способствует активации всех резервов организма для выполнения мышечной работы. Будучи одним из важнейших центров участвует и в регуляции двигательной активности мозжечок должен принимать участие и в регуляции вегетативного обеспечения мышечной активности. Итак мозжечок не только координирует произвольную мышечную активность с процессами поддержания равновесия но и одновременно способствует согласование вегетативных функций в этот период.
Мозжечок влияет на возбудимость вегетативных нервных центров и тем самым способствует адаптации организма к выполнению двигательных актов. Мозжечок можно рассматривать как посредника между вегетативной нервной системой и соматической нервной системой.(мозжечок влияет на деятельность сердца меняет артериальное давления влияет на глубину и частоту дыхания на моторику секрецию всасывание ЖКТ и т.д.).
44. Особенности нейронной организации ретикулярной формации ствола мозга и ее нисходящее влияние на рефлекторную деятельность спинного мозга.
Ретикулярная формация ствола мозга – это структура, идущая в ростральном (к коре) направлении от спинного мозга к таламусу. Помимо участия в обработки сенсорной информации ретикулярная формация выполняет и функцию двигательной системы. Имеется два скопления нейронов ретикулярной формации причастных к этому: это нейроны продолговатого мозга и нейроны моста. Нейроны ретикулярной формации продолговатого мозга веду себя точно так же, как нейроны красного ядра они активируют альфа-мотонейроны сгибателей и тормозят альфа-мотонейроны разгибателей. Нейроны ретикулярной формации моста наоборот действуют как нейроны вестибулярных ядер. Таким оброзом можно условно все двигательные системы ствола мозга разделить на 2 класса: флексорные системы повышающие активность сгибателей – это нейроны красные ядер и ретикулярной формации продолговатого мозга и экстензорные системы нейроны вестибулярного ядер и ретикулярная формация моста.
45. Ретикулярная формация подобно вестибулярным ядрам и нейронам красного ядра, получает информацию от коры мозга (экстрапирамидный путь) и тесно связана с мозжечком: часть информации от мозжечка идет к нейронам продолговатого мозга ( от ядер пробковидного и шаровидного мозжечка ) а от ядер шатра к нейронам локализованным в мосту. Поэтому ретикулярная формация также принимает участие в регуляции позы. Ретикулярная формация коллектор неспецифического сенсорного потока она возможно на основе этой информации участвует в регуляции мышечной активности. Итак ствол мозга обеспечивает поддержание равновесия в условиях покоя и ускорения. Это проявляется наличии статического и стато-кинетического рефлекса.
46. Гипоталамус является одним из важнейших образований мозга участвующих в регуляции активности нейронов парасимпатической и симпатической нервной системы. Гипоталамус представлен собой скопление более чем 32 пар ядер. Существует большое разнообразие в классификации ядер гипоталамуса. Более академично очевидно деление гипоталамических ядер по их топографическому признаку. В настоящиее время весь гипоталамус разделяют на 4 области или группы ядер: 1 преоптическую 2 переднию 3 среднюю 4 заднюю группу ядер. В каждой из этих областей выделяют отдельные ядра. Преоптическая группа: 1 перивентрикулярное 2 медиальное преоптическое 3 латеральное преоптическое. Передняя группа: 1 супраоптическое 2 супрахизматиское 2 паравентикулярное 3 паравентикулярное 4 переднее гипоталамическое ядро. Средняя группа: 1вентромедиальное 2 дорсомедиальное 3 аркуатное 4 латеральное гипоталамическое. Задняя группа: 1 супрамамиллярное 2 премамиллярное 3 медиальное мамилллярное 4 латеральное мамиллярное 5 субталамическое 6 заднее гипоталамическое 7 перифорниатное. В цело в этих ядра локализуются в центры участвующие в вегетативной регуляции а также нейросекреторные нейроны осуществляющие секрецию нейрогормонов и веществ типа либеринов и статинов. Гипоталамус имеет обширное афферентные и эфферентные связи. все рецепторы в конечном доставляют свою информацию и в ядра гипоталамуса. функции гипоталамуса имеются ядра которые преимущественно активируют либо парасимпатические нейроны ствола и спинного мозга, либо симпатические нейроны спинного мозга. Ядра гипоталамуса являются высшими вегетативными центрами. в частности известно что в гипоталамусе имеются центры терморегуляции. Среди ядер передней группы имеются нейроны скопление которые регулируют процесс отдачи тепла поэтому эти скопления называются центром физической терморегуляции. Среди ядер задней группы гипоталамуса имеются скопления нейронов ответственных за процесс теплопродукции это центр химической терморегуляции. В гипоталамусе находятся центры регуляции водного солевого обмена. В гипоталамусе находятся центры белкового углеводного и жирового обмена, центры регуляции сердечно-сосудистой системы, проницаемости сосудов и тканевых мембран , центры регуляции мочеотделения, регуляции сна и бодрствования, полового, эмоционального поведения.
1. Супрахиазматическое ядро – ядро передней группы гипоталамуса. Его нейроны имеют отношения к регуляции полового поведения а также регуляции циркадных ритмов. Показано что это ядро является водителем ритма для пищевого и питьевого поведения для цикла сон – бодрствование , двигательной активности, содержание в крови АКТГ , серотонина, мелатонина , температуры тела. Нейроны супрахизматичесгкого ядра обладают свойствами автоматии и поэтому являются внутренними часами организма. За счет наличия прямых связей этого ядра с сетчаткой глаза ритм нейронов этого ядра приурочен к изменению освещенности в связи с чем многие циркадные ритмы зависят от времени суток. Нейроны этого ядра получают воздействия от центров модулирующих биоритм.
2. Супраптическое и паравентрикурное ядра. Нейроны этих образований помимо участия в процессах регуляции водного- солевого поведения, лактации , активации матки , способы непосредственно продуцировать гормоны пептидной природы – окситоцин , антидиуретический гормон и ряд других. Окситоцин и АДГ по аксонам этих нейронов приходят в составе ножи гипофиза в нейрогипофиз и здесь через аксовазальный синапс выделяется непосредственно в капилляры нейрогипофиза. Нейроны супраоптического и паравентрикулярного ядер помимо секреции этих же гормонов – управляют процессом синаптической передачи , где в качестве медиатора фактически выступает окситоцин и АДГ. Часть нейронов гипоталамуса продуцирует пептидные гормоны , которые получили на звание либеринов и статинов, гормонов которые тормозят выделение других гормонов. Область в которой локализованы эти нейроны называется гипофизарной областью, а нейроны продуцирующие либерины и статины – тубероинфундибулярные нейроны. С током крови либерины и статины достигают передней доли гипофиза и вызывает изменения продукции соответствующего гормона аденогипофиза.
Таким образом благодаря обширной связи гипоталамуса с различными структурами мозга за счет продукции гормонов и нейросекреции гипоталамус участвует в регуляции многих функций организма через гуморальное звено регуляции, изменяя продукцию гормонов гипофиза.
Гипоталамус является компонентом лимбической системы. Гипоталамус является центральной структурой лимбической системы: именно благодаря гипоталамусу все эмоциональные реакции, которые реализуются с участием лимбической системы мозга приобретают конкретную вегетативную и эндокринную окраску.
47. Лимбическая система представляет собой функциональное объединение структур мозга (гиппокамп, миндалина, перегородка и поясная извилина), участвующих в организации эмоционально-мотивационного поведения, таких как пищевой, половой, оборонительный инстинкты. Эта система участвует в организации цикла бодрствование—сон.
Функции лимбической системы:
1. Лимбическая система, как свидетельствуют многочисленные наблюдения, принимает участие в поддержании гомеостаза внутренней среды организма: сохранении относительного постоянства величины артериального давления, газового состава крови, осмотического давления и др.
2. Она осуществляет высшую координацию функций, направленных на поддержание и сохранение индивидуума в постоянно меняющихся условиях окружающей среды, в том числе принимает участие в обеспечении организма питательными веществами и выведение продуктов метаболизма.
3. Лимбическая система принимает участие в осуществлении различных поведенческих реакциях (пищевое, половое поведение, стадная охота животных и др.) и в процессах, связанных с размножением. Эксперименты свидетельствуют, что при хронической стимуляции определенных структур лимбического комплекса наблюдается или гиперсексуализм, или гипосексуализм. При разрушении определенных участков миндалины самки не оберегают потомство от опасности, перестают следить и ухаживать за малышами.
4. Большое значение лимбическая система имеет в формировании памяти. Особая роль в этом приписывается гиппокампу.
5. Лимбико-ретикулярный комплекс принимает активное участие в регуляции гормональной системы. Например, гиппокамп тормозит функцию гипоталамо-гипофизарно-надпочечниковой функции, в то время как миндалевидный комплекс, напротив, ее усиливает.
6. Лимбическая система принимает самое активное участие в формировании различных эмоциональных состояний, создает тот исходный эмоциональный фон, на котором формируется условнорефлекторная деятельность. При электрическом воздействии на различные отделы этой системы в одних случаях наблюдается чрезмерная агрессивность, ярость, в других страх, реакция избегания.
Значимыми образованиями лимбической системы
являются:
1) гиппокамп. Его повреждение ведет к нарушению процесса запоминания, обработки информации, снижению эмоциональной активности, инициативности, замедлению скорости нервных процессов, раздражение — к повышению агрессии, оборонительных реакций, двигательной функции;
2) миндалевидные ядра. Их повреждение ведет к исчезновению страха, неспособности к агрессии, гиперсексуальности, реакций ухода за потомством, раздражение — к парасимпатическому эффекту на дыхательную и сердечно=сосудистую, пищеварительную системы;
3) обонятельная луковица, обонятельный бугорок.
48. Таламус — парное образование, наиболее крупное скопление серого вещества в промежуточном мозге.
Топографически выделяют передние, средние, задние, медиальные и латеральные группы ядер.
По функции выделяют:
1) специфические:
а) переключающие, релейные. Получают первичную информацию от различных рецепторов. Нервный импульс по таламокортикальному тракту идет в строго ограниченную зону коры головного мозга (первичные проекционные зоны), за счет этого возникают специфические ощущения. Ядра вентрабазального комплекса получают импульс от рецепторов кожи, проприорецепторов сухожилий, связок. Импульс направляется в сенсомоторную зону, происходит регуляция ориентировки тела в пространстве;
б) ассоциативные (внутренние) ядра. Первичный импульс идет от релейных ядер, перерабатывается (осуществляется интегративная функция), передается в ассоциативные зоны коры головного мозга;
2) неспецифические ядра. Это неспецифический путь передачи импульсов в кору головного мозга, изменяется частота биопотенциала (моделирующая функция);
3) моторные ядра, участвующие в регуляции двигательной активности.
49. Базальные ядра - это структура переднего мозга: хвостатое ядро, скорлупа чечевицеобразного тела, бледный шар, субталамическое ядро; функционально к этим структрурам относят черную субстанцию среднего мозга.
Функция базальных ганглиев:
Базальные ганглии обеспечивают регуляцию двигательных и вегетативных функций, участвуют в осуществлении интегративных процессов высшей нервной деятельности.
Нарушения в базальных ядрах приводит к моторным дисфункциям, таким как, замедленность движения, изменения мышечного тонуса, непроизвольные движения, тремор. Эти нарушения фиксируются при болезни Паркинсона и болезни Хантингтона.
Получая информацию от ассоциативных зон коры, базальные ядра участвуют в создании программы целенаправленных движений с учетом доминирующей мотивации. Далее соответствующая информация от базальных ядер поступает в передний таламус, где она интегрируется с информацией, приходящей от мозжечка. Из таламических ядер импульсация достигает двигательной коры, которая отвечает за реализацию программы целенаправленного движения через посредство нижележащих стволовых и спинальных двигательных центров.
50. Современные представления о локализации функции в коре головного мозга
В коре головного мозга существуют проекционные зоны.
Первичная проекционная зона - занимает центральную часть ядра мозгового анализатора. Это совокупность наиболее дифференцированных нейронов, в которых происходит высший анализ и синтез информации, там возникают четкие и сложные ощущения. К этим нейронам подходят импульсы по специфическому пути передачи импульсов в коре головного мозга (спиноталамический путь).
Вторичная проекционная зона - расположена вокруг первичной, входит в состав ядра мозгового отдела анализатора и получает импульсы от первичной проекционной зоны. Обеспечивает сложное восприятие. При поражении этой зоны возникает сложное нарушение функции.
Третичная проекционная зона - ассоциативная - это полимодальные нейроны, разбросанные по всей коре головного мозга. К ним поступают импульсы от ассоциативных ядер таламуса и конвергируют импульсы различной модальности. Обеспечивает связи между различными анализаторами и играют роль в формировании условных рефлексов.
Кора большого мозга - поверхностный слой полушарий большого мозга, образованный его серым веществом; высший отдел центральной нервной системы. Кора: - обеспечивает высшую нервную деятельность человека; - способствует адаптации организма в меняющихся условиях среды; - регулирует приобретенные формы поведения; и - обеспечивает уровень психических способностей человека.
На основе данных филогенеза в коре большого мозга выделяют: - древнюю и старую кору, которые играют решающую роль в регуляции вегетативных функций, инстинктивном поведении, возникновении эмоций; и - новую кору, разные участки которой отвечают за выполнение моторных, сенсорных и ассоциативных функций.
Пластичность коры головного мозга - способность корковых клеток замещать функцию выпавших участков.
Основные функции коры больших полушарий:
1) интеграция (мышление, сознание, речь);
2) обеспечение связи организма с внешней средой, приспособление его к ее изменениям;
3) уточнение взаимодействия между организмом и системами внутри организма;
4) координация движений.
Эти функции обеспечиваются корригирующими, запускающими, интегративными механизмами
Физиологические основы психических функций человека
12. Понятия о высших функциях человека, их адаптивная роль. Психика - это специфическое свойство высокоорганизованной материи -головного мозга, заключающееся в отражении предметов и явлений существующего вне нас и независимо от нас материального мира. Будучи особым свойством высокоорганизованной материи психика прошла долгий путь развития от самых элементарных форм формирования ощущений до сознания человека. Ощущения и восприятия являются необходимым начальным этапом наших знаний о самом себе и о внешнем мире.
0щущение_- это процесс отражения в ЦНС отдельных свойств предметов и явлений объективной реальности, непосредственно воздействующей на органы чувств. Всякое ощущение имеет качество, силу, длительность. Качественные особенности тех или иных ощущений называются модальностью.
Являясь простейшей первоначальной формой отражения материального мира в нашем сознании, наши ощущения дают материал для более сложных форм отражения действительности в сознании человека - для восприятия, представления, для разных форм мысли. Поэтому ощущения являются источником всех знаний об окружающей нас действительности. Процесс возникновения ощущений представляет собой не что иное, как превращения энергии внешнего раздражения в факт сознания. Этот процесс неразрывно связан с деятельностью ЦНС.
Все ощущения можно разделить на две группы.
1. Ощущения, отражающие свойства предметов и явлений материального мира. К ним относятся: зрительные, вкусовые, слуховые, обонятельные и кожные.
2. Ощущения, отражающие движения отдельных частей нашего тела и состояние внутренних органов. К ним относятся: двигательные ощущения, ощущения равновесия и др.
Восприятие, в основе которого лежат ощущения, представляет собой сложный процесс приема и преобразования информации, обеспечивающей организму отражение объективной реальности и ориентировки в окружающем мире. Восприятие - это активный процесс, характеризующийся направленностью и избирательностью, т. е. выделением из массы разнородных объектов внешнего мира именно тех, которые более всего необходимы именно в данный момент.
Ощущения и восприятия человека носят сознательный, осмысленный характер. Важную роль в этом играют мышление и речь.
Мышление представляет собой высшую форму отражения мира. Мышление позволяет получить знания о таких объектах, свойствах и сношениях реального мира которые не могут быть непосредственно восприняты на чувственной ступени познания. Мышление как высшая и качественно новая форма психической деятельности возникло на базе общественного развития, благодаря труду.
Мышление - процесс опосредованного обобщения отражения действительности с ее связями, отражениями и закономерностями. С помощью мышления познается содержание, значение и смысл воспринимаемого, а также внутренние, скрытые для непосредственного познания (ощущения и чувственного восприятия) особенности предметов и явлений. С помощью мышления человек может понять не только настоящее, но и будущее и прошедшее, строить гипотезы, обобщать и обеспечивать их проверку. Наиболее высокая форма обобщения - понятие, наивысшая форма мышления - понятийное мышление. В понятии отражено то, что не может непосредственно быть воспринято при помощи наших органов чувств, причем, понятие отражает в предмете лишь существенное.
Характерной, отличительной особенностью человеческого мышления является его неразрывная связь с речью, языком.
Язык - это средство выражения мысли и формы существования мысли. Человеческий язык представляет систему материальных средств: звуков, слов, форм слов и закономерных сочетаний слов в предложении. Язык теснейшим образом связан с мышлением, он регистрирует и закрепляет в словах и предложениях результаты работы мышления, успехи познавательной деятельности человека и таким образом делает возможным обмен мыслями в человеческом обществе. Без языка человеческая мысль не могла бы существовать и развиваться. Одной из важнейших специфических особенностей человеческого познания является речь, язык, который дает возможность создавать научные понятия, категории, формулировать законы науки и тем самым глубоко проникать в сущность явлений.
13. Особенности восприятия у человека. Внимание. Значение работ И. П. Павлова и А. А. Ухтомского для понимания физиологических механизмов внимания. Физиологические корреляты внимания.
Внимание - это сосредоточенность и направленность психической деятельности на каком-то определенном объекте. С помощью внимания обеспечивается отбор необходимой информации. Различают непроизвольную и произвольную формы внимания. Непроизвольное внимание - это первичный врожденный процесс, который осуществляется с помощью наследственных нервных механизмов. Произвольное внимание выражается в целенаправленной мобилизации психической активности человека, производится на сознательно преднамеренно выбранный объект внимания. Произвольное внимание свойственно человеку и возникло в процессе трудовой деятельности. Характеристиками внимания являются: устойчивость, переключаемость, объем, выражающийся в количестве объектов, которые могут быть восприняты и
запечатлены человеком в относительно короткий период времени.
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ВНИМАНИЯ связаны с проявлениями ориентировочного рефлекса, с локальными процессами активации коры головного мозга, а также с эмоциональными состояниями и биологическими потребностями организма. В связи с тем, что различные стадии внимания от диффузионного до высококонцентрированного коррелируют с определенными, различными по уровню, состояниями бодрствования, можно предположить наличие некоторых общих нейрофизиологических механизмов внимания и бодрствования. Избирательное внимание возможно лишь при состоянии повышенного, но не чрезмерно высокого бодрствования, которое на ЭЭГ характеризуется высокочастотными десинхронизированными низкоамплитудными колебаниями с редким появлением групп синхронизированных альфа-волн. Диффузным формам внимания соответствуют состояния расслабленного бодрствования с четко выраженными на ЭЭГ синхронизированными альфа-ритмами. Рассеянное внимание, отвлекаемость, трудность сосредоточения часто возникают в состоянии сильного возбуждения, связанного с сильными эмоциями, при этом на ЭЭГ регистрируется десинхронизированная высокочастотная низкоамплитудная электрическая активность.
Большинство исследователей считает, что анатомическим субстратом регуляции различных состояний бодрствования, форм внимания и сознания является ретикулярная формация различных отделов ствола мозга, диффузная таламическая система с ее корковыми проекциями, субталамус и гипоталамус. Неспецифическая таламическая система рассматривается как фильтрующий механизм, обладающий способностью обострять и переключать внимание с одних раздражителей на другие.
Ассоциативные зоны коры являются центральным звеном в системе механизмов, регулирующих селективный отбор информации, собирательные формы восприятия, внимания и сознания.
С работой кортико-ретикулярных механизмов связывают в основном процессы произвольного внимания, которому в отличие от непроизвольного, предшествует подготовительная установка, соответствующая концентрации внимания. Оно представляет собой подготовительное состояние для специфической деятельности. Непроизвольное внимание может осуществляться на подкорковом уровне, произвольное внимание возникает при действии выработавшихся в процессе обучения условных стимулов и требует участия корково-ретикулярных механизмов. Выделяют особую форму произвольного внимания - интеллектуальное внимание, объектом которого являются мысли и воспоминания. Ведущая роль в осуществлении этого высшего вида внимания принадлежит фронтальным кортико-фугальным влияниям на различные области и центры мозга.
Физиологические механизмы, регулирующие разные формы внимания, функционируют как колебательная система, работающая по принципу градуальных изменении. В избирательном сенсорном внимании, а также в интеллектуальном внимании особое значение придается корковым фронтальным областям по-видимому, обеспечивающих и речевую регуляцию внимания.
14. Память и ее значение в формировании целостных приспособительных реакций.
Память - одно из основных свойств ЦНС, выражающееся в способности на короткое или длительное время сохранять информацию (отпечатки, следы) о событиях внешнего мира и реакциях организма. Память как единый процесс складывается из трех взаимосвязанных этапов: запоминания, хранения опыта и воспроизведения опыта.
ВИДЫ ПАМЯТИ. Различают память наследуемую (генетическую) и ненаследуемую (индивидуальную). Кроме того, выделяют и такие виды памяти, как образная (которая воспроизводит образ жизненно важного объекта), эмоциональную (когда аналогичная ситуация вызывает эмоции, характерные для происходивших ранее в_ ситуации событий, словесно-логическую (она обусловлена развитием речи и свойственна только человеку). По времени сохранения информации различают: непосредственный отпечаток сенсорной информации (сенсорная память), кратковременную и долговременную память.
Непосредственный отпечаток обеспечивает удержание следов в сенсорной памяти не более 500 мс. Сенсорная память человека не зависит от его воли и не может быть подвергнута сознательному контролю. Этот вид памяти зависит от функционального состояния организма и обладает индивидуальными особенностями. Время сохранения образа внешнего мира неодинаково для различных органов чувств. Наиболее длительно сохраняются зрительные образы. Непосредственный отпечаток сенсорной информации невоспроизводим. Он является начальным этапом переработки поступивших сигналов. Количество информации, содержащейся в образе, "отпечатке", избыточно. В дальнейшем высший аппарат анализа информации определяет, вычленяет и интегрирует лишь существенную ее часть.
Кратковременная память формируется на базе непосредственного отпечатка сенсорной информации. Обеспечивает удержание ограниченной части поступающих сигналов из внешней среды, позволяет воспроизводить какую-то часть предъявляемого материала и тем самым некоторое время использовать определенное количество информации.
Долговременная память обеспечивает сохранение информации неограниченное время. В системе долговременной памяти, объем которой практически не ограничен, сохраняется огромное количество информации без ее искажения. Информация при необходимости может легко воспроизводиться.
ПРОЦЕССЫ ПАМЯТИ. Процесс эапечатления в ЦНС поступающей информации может быть двух видов: произвольным и непроизвольным. Произвольное запечатление оказывается более эффективным. Избыток зафиксированной информации не способствует успешной переработке и ее дальнейшему использованию. Для каждого индивида существует оптимальный объем информации, который может быть успешно переработан и освоен. Стимулы, имеющие большое биологическое и социальное значение, фиксируются значительно эффективнее независимо от их физической силы.
Сохранение следов является центральным звеном в системе памяти. В процессе накопления и хранения приобретенной информации в ЦНС происходит ее сложная переработка. Неоднократное повторение идентичных или подобных воздействий запечатлевается в памяти не по принципу простого наложения и закрепления, а путем .упорядочения и обогащения.
Воспроизведение заключается в извлечении (считывания) информации из систем или блоков памяти. Воспроизведение, как и запоминание, может быть произвольным и непроизвольным. Непроизвольное воспроизведение происходит непреднамеренно и может иметь навязчивый характер. Произвольное воспроизведение, заключающееся в воспроизведении из долговременной памяти ранее приобретенной информации, имеет избирательный характер и представляет собой активный процесс, требующий включения внимания, а иногда и значительных умственных усилий.
Под забыванием понимают невозможность воспроизведения , неиспользование и неприменение в практической деятельности приобретенной информации, которая тем не менее при определенных обстоятельствах может воспроизводиться.
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ КРАТКОВРЕМЕННОЙ ПАМЯТИ. Существуют несколько теорий, рассматривающих физиологические механизмы кратковременной памяти. На основании морфологических данных, свидетельствующих о существовании замкнутых цепейнейронов была предложена теория реверберации импульсов по замкнутой системе нейронов. Согласно этой теории, субстратом, хранящим поступающую информацию, является так называемая нейронная ловушка, образующаяся из цепи нейронов, что обеспечивает длительную циркуляцию возбуждения по таким кольцевым связям. Если импульсация, подобная той, которая сформировала реверберационную цепочку, будет повторно поступать к тому же нейрону, то возникает закрепление следов этих процессов в памяти. Отсутствие повторной импульсации или приход тормозного импульса к одному из нейронов цепочки реверберации, приводит к прекращению реверберации, забыванию.
Электротоническая теория памяти основана на том, что кратковременная память может быть объяснена специфическими явлениями, развивающимися при прохождении нервных импульсов через синапсы и развитии в них электротонических потенциалов, которые регистрируются в течение нескольких минут и даже часов и способны облегчить прохождение имульсов через строго определенные синапсы. Сильное раздражение нейронов любого уровня ЦНС часто приводит к явлению посттетанической потенциации, которая выражается в нарастании возбудимости этого нейрона и развитии длительной импульсной активности после прекращения раздражения.
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ. Этот вид памяти не может базироваться только на циркуляции импульсов или изменениях электрофизиологических характеристик отдельных нейронов. При различных воздействиях на организм (гипоксия, наркоз, охлаждение, сон) могут разрушаться кольцевые реверберационные связи и снижаться возбудимость нейронов. При этом огромное количество информации сохраняется в долговременной памяти в неизмененном виде.
Согласно анатоической теории запоминание и хранение информации осуществляется за счет образования новых терминальных волокон, изменения их формы, размеров, развития шипикового аппарата на дендритах нейронов, устанавливающих связи с другими нейронами.
Глиальная теория основывается на изменениях глиальных клеток, которые окружают нейроны. Глиальные клетки могут синтезировать особые вещества, облегчающие синаптическую передачу или повышающие возбудимость соответствующих нейронов.
Биохимическая теория развивает представление об активировании ферментативных процессов при образовании медиаторов или перестройке мембраны нейронов. Было обнаружено, что при активации нейрональных процессов в них происходит интенсификация белкового обмена. Исследованиями показано, что в хранении и воспроизведении следов информации необходимы специфические белки. Кроме того, показано, что торможение синтеза белка приводит к нарушению или прекращению консолидации следов в долговременной памяти. Уже можно с уверенностью сказать, что в механизмах долговременной памяти перестройка структур молекул ДНК и РНК в нейронах головного мозга играет первостепенную роль.
15. Биологическая роль эмоций. Теории эмоций. Вегетативные и соматические компоненты эмоций. Виды и роль эмоций в возникновении психосоматических заболеваний у человека.
Эмоции. Одним из проявлений ВНД являются эмоции. Они представляют собой реакции организма, имеющие ярко выраженную субъективную окраску, на воздействия внешних и внутренних раздражителей. С помощью эмоций определяется личностное отношение человека к окружающему миру и к самому себе. Эмоции - один из ведущих механизмов регуляции адаптивной и психической деятельности организма. Эмоциональные состояния реализуются в определенных поведенческих реакциях.
Эмоции возникают на этапе оценки вероятности удовлетворения или неудовлетворения возникших потребностей, а также при удовлетворении этих потребностей.
Биологическое значение эмоций состоит в выполнении ими сигнальной и регуляторной функции. Сигнальная функция эмоций заключается в том, что они сигнализируют о полезности или вредности данного воздействия, успешности или неуспешности выполняемого действия. Приспособительная роль этого механизма заключается в немедленной реакции на внезапное воздействие внешнего раздражения, поскольку эмоциональное состояние мгновенно вызывает ярко выраженные переживания определенной окраски. Это приводит к быстрой мобилизации всех систем организма к осуществлению ответной реакции, характер которой зависит от того, сигналом полезного или вредного воздействия на организм служит данный раздражитель. Таким образом, воздействия, исходящие как из внешней среды, так и от самого организма, приводят к возникновению эмоциональных переживаний, дающих общую качественную характеристику воздействующему фактору, опережая его более полное, детальное восприятие.
Регуляторная функция эмоций проявляется в формировании активности, направленной на удовлетворение возникших потребностей, также на прекращение действия раздражителей, т.е. в реализации механизмов адаптации организма к непрерывно меняющимся условиям среды. Неудовлетворенные потребности организма обычно сопровождаются эмоцией, носящей неприятный характер. Удовлетворение исходной потребности, как правило, сопровождается приятным эмоциональным переживанием.
Возникновение положительных эмоций при удовлетворении потребности характеризует успех поиска для достижения цели, что ведет к прекращению дальнейшей поисковой деятельности. С другой стороны, неоднократное удовлетворение тех или иных потребностей, сопровождающееся субъективно приятными эмоциональными переживаниями, приводит к тому, что в дальнейшем организм стимулируется к целенаправленной деятельности представлением о будущей положительной эмоции. Этот второй стимулирующий момент целенаправленного мотивационного поведения - результат общения, и поэтому приобретает особое значение в дальнейшем поведении человека и животного.
Эмоции делятся на положительные и отрицательные. Положительные эмоции определяют такое состояние организма, которое характеризуется активными усилиями животного и человека, направленными на сохранение и усиление этого состояния. Отрицательные эмоции проявляются в усилиях, направленных на устранение неблагоприятного состояния организма, вызванного неудовлетворением потребности или воздействием вредоносного фактора. Положительные и отрицательные эмоции играют существенную роль в приспособительном поведении.
Эмоции также делятся на низшие и высшие. Низшие эмоции более элементарны, связаны с органическими потребностями животных и человека и подразделяются на два вида:
* гомеостатические, направленные на поддержание гомеостаза организма и носящие всегда отрицательный характер;
* инстинктивные, связанные с половым инстинктом, инстинктом сохранения рода и другими поведенческими реакциями.
Высшие эмоции возникают только у человека в связи с удовлетворением социальных и идеальных потребностей (интеллекуальных, моральных, эстетических и др.). Эти более сложные эмоции развивались на базе сознания и оказывают контролирующее и тормозящее влияние на низшие эмоции.
Различают эмоции стенические - вызывающие активную деятельность и астенические - снижающие активность. По длительности и степени выраженности различают эмоции настроения, страсти, аффекта.
НЕРВНЫЙ СУБСТРАТ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ ЭМОЦИЙ. Экспериментально было показано, что эмоции гнева и страха под влиянием таламических разрядов сопровождаются повышенным поступлением адреналина в кровь, что приводит к развитию симпатикотонии, играющей роль в подготовке организма к деятельности. Эта теория получила название таламической теории эмоций. В норме положительные эмоции умеренной интенсивности связаны преимущественно с парасимпатическими реакциями. Отрицательные эмоции - с симпатическими и включают в большой степени симпато-адреналовую и в меньшей ваго-инсулиновую системы. При сильном эмоциональном возбуждении нисходящие гипоталамические влияния не ограничиваются одним из отделов вегетативной нервной системы.
В соответствии с теорией И. Пейпеца возникновение эмоций обусловливается лимбической системой. Кортикальные эмоциональные процессы возникают в гиппокампе, оттуда импульсы идут в маммилярные тела, затем в передние ядра гипоталамуса и в поясную извилину (круг Пейпеца). Эмоциональная окраска психических процессов создается распространением этих импульсов на другие области коры. Рецептивной областью эмоциональных переживаний является поясная извилина. Целостность этой цепи предоставляет собой механизм, организующий переживание и выражение эмоций. Эмоции возникают либо сначала в коре, откуда импульсы поступают в "круг" через гипокамп, либо в результате возбуждения гипоталамуса , и тогда кору поясной извилины следует рассматривать как воспринимающую область для эмоциональных ощущений в результате приходящих из гипоталамуса импульсов. Лимбическую систему рассматривают как "висцеральный мозг". Считают, что именно здесь происходит интеграция воспринимаемой информации, получаемой от всех структур тела, в том числе от скелетной мускулатуры и внутренних органов, и формирование определенных эмоциональных состояний.
В настоящее время принято считать, что нервным субстратом эмоций является лимбико -гипоталамический комплекс. Включение гипоталамуса в эту систему обусловлено тем, что множественные связи гипоталамуса с различными структурами головного мозга создают физиологическую и анатомическую основу для возникновения эмоций.
Новая кора на основе взаимодействия с другими структурами, особенно гипоталамусом, лимбической и ретикулярной системами, а также между различными областями самой новой коры, несомненно играет важную роль в субъективной оценке эмоциональных состояний.
Эмоциональные состояния являются важной формой адаптационных состояний организма, и играют огромную роль в создании условий для более широкого и более совершенного приспособления животных и человека к окружающим условиям.
Механизм возникновения эмоций может быть рассмотрен с точки зрения биологической теории П. К. Анохина. Суть этой теории заключается в том, что положительные эмоции при удовлетворении какой-либо потребности возникают только в том случае, когда параметры реально полученного результата точнейшим образом совпадают с параметрами предполагаемого результата, запрограммированного в акцепторе результатов действия. В таком случае возникает реакция согласования, которая субъективно сопровождается чувством удовлетворения, положительными эмоциями. Если параметры реально полученного результата не совпадают с запрограммированными в акцепторе результатов действия, это сопровождается чувством неудовлетворения, беспокойства - отрицательными эмоциями. Это приводит к формированию ориентировочно-исследовательской реакции и к формированию новой комбинации эффекторных возбуждений, необходимых для организации нового полноценного периферического акта, который обеспечит получение результата, параметры которого совпадают с запрограммированными в акцепторе результатов действия.
16. Роль эмоций в целенаправленной деятельности человека. Эмоциональное напряжение (эмоциональный стресс) и его роль в формировании психосоматичиских заболеваний организма.