

М.Д. НАРІЙЧУК, Т.А. РЕШЕТНЯК
МЕДИЧНА БІОЛОГІЯ
За редакцією професора А.Д. Тимченка
ЗАТВЕРДЖЕНО Департаментом кадрової політики, освіти і науки МОЗ України як підручник для студентів вищих медичних навчальних закладів І—III рівнів акредитації
Бібліотека Бердянською медичного коледжу
КИЇВ «МЕДИЦИНА» 2009
Резензенти: ВД. Кірсей, директор, ОМ.Андрющенко, голова циклової комісії фундаментальних та професійно-орієнтованих дисциплін Білгород-Дністровського медичного училища ГД. Бердишев, проф. Київського Національного університету
імені Тараса Шевченка
Н38 Медична біологія: підручник / М.Д. Нарійчук, Т.А. Решетняк; за ред. проф. А.Д. Тимченка. — К.: Медицина, 2009. — 264 с.
І8ВК 978-966-10-0069-7
Підручник підготовлено відповідно до програми з медичної біології. Вміщено інформацію, необхідну для практичної діяльності сучасного працівника охорони здоров'я. Зокрема розглянуто основи генетики людини, методи вивчення спадковості, спадкові хвороби людини. В окремих розділах висвітлено питання медичної протозоології, медичної гельмінтології та арахноен-томології.
Для студентів вищих медичних навчальних закладів І—III рівнів акредитації.
ББК 56.13я73
© М.Д. Нарійчук, Т.А. Решетняк, 2009
I8ВN 978-966-10-0069-7 ©Видавництво «Медицина» 2009
ЗМІСТ
вступ
РОЗДІЛ 1. Структурно-функціональна організація клітини.
Розмноження на клітинному рівні .9
Структурно-функціональна організація еукаріотичної клітини .9
Морфологія клітини. Органели загального і спеціального призначення..............................................15
Хімічний склад клітини . ..22
Життєвий цикл клітини. Поділ клітини ..........39
РОЗДІЛ 2. Молекулярні основи спадковості. Реалізація спадкової інформації ................................47
Характеристика нуклеїнових кислот: ДНК і РНК, просторова організація, видова специфічність, роль у зберіганні та передачі спадкової інформації ............................................................................47
Реплікація ДНК ..53
Підтримування генетичної стабільності клітин: самокорекція і репарація ДНК.................................57
Будова гена прокаріотів та еукаріотів ..60
Генетичний код, його властивості ..64
Біосинтез білків ..67
Екзонно-інтронна організація генома еукаріотів ..73
Генна інженерія. Біотехнологія ..74
РОЗДІЛ 3. Закономірності успадкування менделівських ознак
у людини. Взаємодія генів .81
Генетика: предмет і завдання, етапи розвитку, основні
терміни і поняття .81
Моногібридне схрещування: закон одноманітності
гібридів першого покоління, закон розщеплення .87
Взаємодія генів .91
Полігенне успадкування ознак у людини .94
Плейотропна дія гена .96
РОЗДІЛ 4. Хромосомна теорія спадковості. Генетика статі .97
Зчеплене успадковування. Механізм кросинговеру .97
Хромосомна теорія спадковості .99
Генетичні карти хромосом .100
Генетика статі .....100
Успадковування захворювань людини, зчеплених зі статтю .........102
РОЗДІЛ 5. Мінливість у людини як властивість життя
і генетичне явище ..105
Мінливість, її форми та прояви на рівні організму: фенотипова,
генотипова мінливість. Модифікації та норма реакції .105
Роль генотипу і навколишнього середовища в мінливості ознак .106
Комбінативна мінливість. Мутаційна мінливість .107
Класифікація мутацій .107
Мутагени: фізичні, хімічні, біологічні .109
Генетична небезпека забруднення середовища .110
Закон гомологічних рядів спадкової мінливості, його
практичне значення .112
РОЗДІЛ 6. Основи генетики людини.
Методи вивчення спадковості .114
Основи медичної генетики. Людина як специфічний об'єкт
генетичного аналізу. Методи вивчення спадковості людини 114
Генеалогічний метод. Правила побудови родоводів. Генетичний
аналіз родоводів 116
Близнюковий метод 124
Визначення взаємодії генотипу та довкілля в прояві
патологічних ознак людини 126
Цитогенетичний, біохімічний методи та ДНК-діагностика 127
^ Метод дерматогліфіки ....133
Метод гібридизації соматичних клітин 135
РОЗДІЛ 7. Спадкові хвороби людини .137
Класифікація спадкових хвороб людини 137
Хромосомні хвороби, спричинені порушенням кількості
або структури хромосом 138
Моногенні (молекулярні) спадкові захворювання 145
Молекулярні хвороби вуглеводного, амінокислотного,
ліпідного та мінерального обміну 146
Пренатальна діагностика спадкової патології 155
РОЗДІЛ 8. Біологія індивідуального розвитку. Молекулярно-генетичні механізми онтогенезу.
Патологічні порушення онтогенезу людини 158
Гаметогенез................. 158
За п л ід нення........................................................................................................................................161
Онтогенез: типи, періоди, етапи 161
Етапи ембріонального розвитку людини 163
Диференціювання на молекулярно-генетичному, клітинному
та тканинному рівнях 165
Природжені вади розвитку 166
Ембріональна індукція 170
Регуляція в процесі дроблення та її порушення (близнюки,
вади розвитку, виродливість) 173
Періоди постембріонального розвитку людини 175
Особливості постнатального періоду індивідуального розвитку
людини у зв'язку з її біосоціальною суттю ....179
Біополе, біологічні ритми та їхнє медичне значення ....181
Види та шляхи регенерації ....183
Види трансплантації тканин у людини ....187
Старість як завершальний етап онтогенезу людини.
Теорія старіння ....188
РОЗДІЛ 9. Медико-біологічні основи паразитизму. Медична
протозоологія. Найпростіші паразити людини 191
Вступ до медичної паразитології. Походження та еволюція
паразитизму 191
Принципи класифікації паразитів 193
Принципи взаємодії паразита і хазяїна 194
Характерні риси і класифікація підцарства Найпростіші (Ргоіогоа)............................ ...196
РОЗДІЛ 10. Медична гельмінтологія. Плоскі та Круглі черви —
паразити людини 209
Тип Плоскі черви (Ріаіпеїтіпіпев) 209
Тип Круглі черви (№таІпе1тіпІЇіе8) 220
РОЗДІЛ 11. Медична арахноентомологія 228
Клас павукоподібні (АгасЬпоіаеа) 229
Клас Комахи (Іпзесіа) 232
РОЗДІЛ 12. Взаємозв'язок індивідуального та історичного
розвитку. Біосфера та людина 238
Структура та функції біосфери 238
Основні положення вчення В.І. Вернадського про
організацію біосфери 241
Сучасні концепції біосфери 242
Ноосфера 244
Екологія людини 245
Фактори середовища 246
Єдність організму та середовища 247
Класифікація симбіологічних відносин за природою
взаємовідносин 248
Види екосистем 250
Проникнення людини в біогеоценози 251
Формування антропоценозів 251
Екологічне прогнозування 252
Адаптація людей до екстремальних умов середовища 253
Вплив антропогенних факторів забруднення довкілля
на здоров'я населення 257
Характеристика отруйних для людини рослин і тварин 259
ВСТУП
Біологія (від грец. bios — життя, Іоgos — вчення) — наука про життя. Цей термін уперше вживає в 1794 р. німецький професор анатомії Т. Руз. У 1802 р. Ж. Ламарк і Г. Тревірантус (незалежно один від одного) запропонували його як науковий термін.
Сучасна біологія — це система наук про живу природу, морфологію та функцію живих істот, їх походження, розвиток, поширення, взаємозв'язок між собою та неживою природою.
Медична біологія — це галузь біології, яка вивчає людину як біосоціальний об'єкт, а також різні організми і процеси, які впливають на людину, її життєдіяльність, фізичний та інтелектуальний розвиток. Медична біологія слугує збереженню здоров'я людини, її генофонду. Для цього медична біологія вивчає закономірності спадковості, індивідуального та еволюційного розвитку, морфологічної і соціальної адаптації людини до умов навколишнього середовища.
Знання медичної біології необхідні для вивчення нормальної і патологічної анатомії, фізіології, гістології, мікробіології, медичної генетики, екології та всіх клінічних дисциплін, які потребують медико-біологічної підготовки. Медична біологія дає студентам теоретичну підготовку, необхідну для практичної діяльності сучасного працівника охорони здоров'я.
Медична біологія як самостійна галузь біології вивчає найскладніший життєвий об'єкт на Землі — людину. Ця складність пов'язана з тим, що в людини крім загально біологічних властивостей, притаманних усім живим організмам, є чимало того, що вважається результатом належності її до людського суспільства.
Суспільство створено людиною для можливості виконання певної праці, а співіснування у суспільстві змінило людину як біологічну істоту. Якщо вважати середовищем існування людини суспільство, крім зовнішнього середовища, яке об'єднує всі живі
організми на Землі, людина є біосоціальною істотою. Тому в результаті розвитку суспільства людина має складні психофізіологічні реакції, які безумовно впливають на фізіологічні процеси. Наприклад, добре відомо, що сміх покращує стан людини, а сльози навпаки — погіршують його.
Рівні організації живого
Однією з основних властивостей життя є впорядкованість. Дослідження ступеня вираження впорядкованості є результатом розвитку різних наук (фізики, хімії, біології). Поступово з'являються поняття про рівні організації живого. Рівні організації живого та ступені організації живого наведено в табл. 1.
Таблиця 1. Рівні та ступені організації живого
Ступінь організації живого |
Рівень організації живого |
Біологічні мікросистеми * |
Молекулярний |
|
Субклітинний |
|
Клітинний |
Біологічні мезосистеми |
Тканинний |
|
Органний |
Організмовий |
|
Біологічні макросистеми |
Популяційно-видовий |
Біогеоценозний |
|
|
|
|
Біосферний |
Виділяють три типи зв'язку між системами: ієрархічний, функціональний, сітковий.
Молекулярний рівень — доводить існування різновидів (за молекулярним складом) різних клітин. Але значна частина їх повторюється у багатьох організмів (наприклад, однакові в усіх клітин молекули амінокислот, гліцерину та ін.). Слід вважати, що саме на цьому рівні посилюється диференціація та спеціалізація клітин, а також забезпечується здійснення всіх найважливіших процесів життєдіяльності: обмін речовин, кодування і передавання спадкової інформації.
Клітинний рівень — об'єднує всіх живих істот, для яких клітина — це елементарно-генетична і структурно-функціональна одиниця. Клітина здатна до саморегуляції, самооновлення, розвитку, поділу. Клітина еволюціонувала від прокаріотичної до еукаріотичної. Складні взаємозв'язки між про- та еукаріотичною клітиною збережені і простежуються, наприклад при інфекційних захворюваннях (туберкульоз, черевний тиф та ін.), спричинених певними бактеріями, тобто прокаріотичними клітинами. Якщо організм одноклітинний, то у нього клітинний рівень збігається з організмовим. Це характерно для: туберкульозної палички, пневмокока, пневмотифозної палички та ін.
Тканинний рівень — дає змогу виділити та характеризувати основні типи тканин у рослин і тварин. Тканина — це сукупність клітин, що мають спільне походження, морфологію, функції, та міжклітинної речовини, яка характеризується високою спеціалізацією. Клітини кожної тканини мають певну форму, розміри. На тканинному рівні прослідковується подібність між усіма організмами. Більшість авторів виділяють 5 основних типів тканин рослин: твірна, основна, провідна, покривна, механічна. У тварин 4 основних типи: епітеліальна, сполучна, м'язова, нервова. Морфо-функціональна цілісність рослинного або тваринного організму досягається під час взаємодії та узгодженого розвитку всіх тканин.
Органний рівень — супроводжується функціональною взаємодією кількох типів тканин.
Організмовий (або онтогенетичний) рівень супроводжується взаємодією різних систем органів. На цьому рівні вивчають адаптивні зміни та поведінку організмів у різних екологічних умовах. Крім того, визначено, що вид складається з особин, які при значній подібності мають багато різновидів.
Популяційно-видовий рівень — вивчає популяцію, в якій здійснюються елементарні еволюційні перетворення. Вивчення складу, динаміки популяції є предметом генетики, морфології, фенології, екології та ін.
Біогеоценозний рівень — взаємодіюча сукупність організмів різного рангу з різними чинниками середовища існування.
Біосферний рівень — забезпечує речовинно-енергетичні кру-гообіги. Біосфера як одна з географічних оболонок Землі є єдиним цілим і має свої закономірності. Саме наш співвітчизник академік В.І. Вернадський є автором вчення про біосферу.
РОЗДІЛ 1
Структурно-функціональна організація клітини. Розмноження на клітинному рівні
Структурно-функціональна організація еукаріотичної клітини
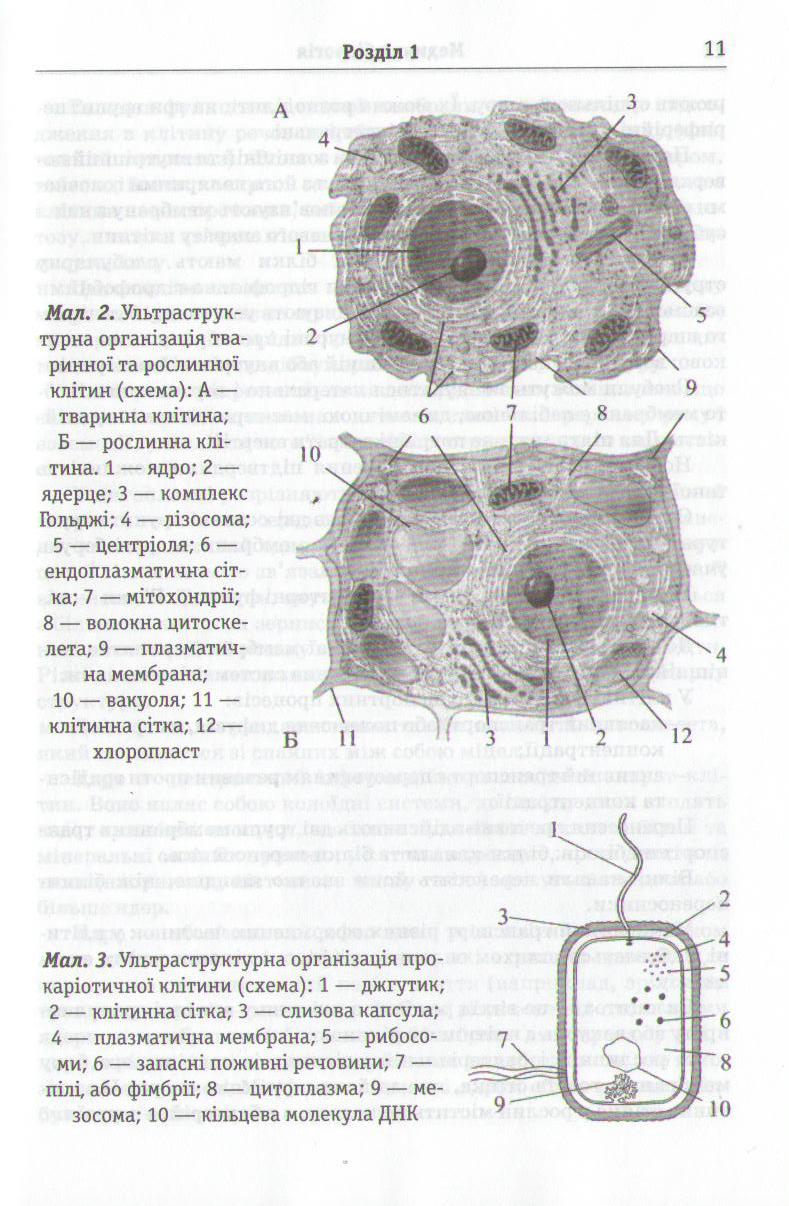
Клітина — основна структурно-функціональна одиниця живих організмів. Клітина найпростіших (амеби, інфузорії, балан-тидії та ін.) перебуває в безпосередній взаємодії з навколишнім середовищем. Клітина найпростіших — це самостійний організм. Клітини рослинних і тваринних організмів мають схожу будову (мал. 1—3).
Вони складаються з плазматичної мембрани, цитоплазми, ядра та органел загального призначення і спеціальних органел.
Клітинні органели поділяють на мембранні та немембранні: мембранні органели мають одну або дві мембрани. До одномемб-ранних органел належать:
1) ендоплазматична сітка:
а) шорстка (гранулярна);
б) гладенька (агранулярна);
пластинчастий комплекс (комплекс Гольджі);
лізосоми;
вакуолі.
До двомембранних:
1) ядро;
2) имітохондрії;
3) пластиди.
А до немембранних органел:
рибосоми;
центріолі;
мікротрубочки;
мікрофіламенти.
Клітинна плазматична мембрана — постійний універсальний компонент поверхневого апарату всіх клітин. Ще в 1925 р. Портер і Грендель, вивчаючи ліпіди оболонок еритроцитів, довели,
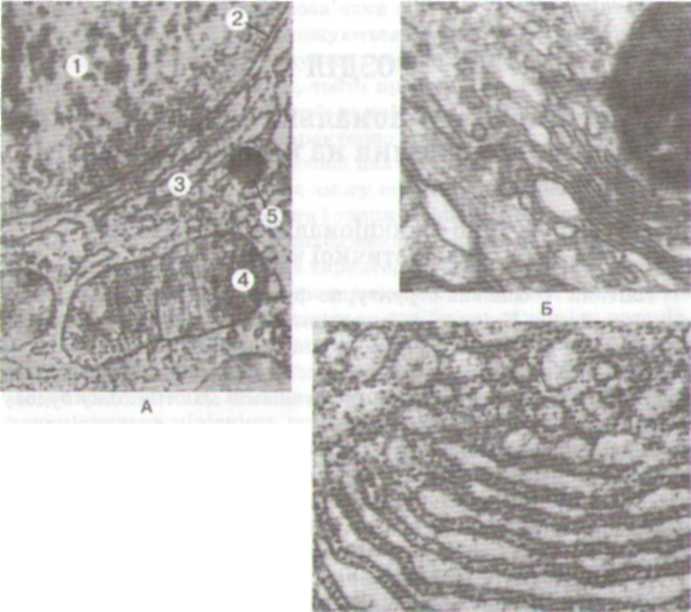 Мал.
1. Електронограми
тваринних
клітин:
А
—
фрагмент
тваринної
клітини:
1
—
ядро;
2
—
ядерна
оболонка;
3
—
ендоплазматичний
ретикулум;
4
—
мітохондрія;
5
—
лізосома.
Б
—
комплекс
Гольджі.
В
—
гранулярна
ендоплазматична
сітка.
Мал.
1. Електронограми
тваринних
клітин:
А
—
фрагмент
тваринної
клітини:
1
—
ядро;
2
—
ядерна
оболонка;
3
—
ендоплазматичний
ретикулум;
4
—
мітохондрія;
5
—
лізосома.
Б
—
комплекс
Гольджі.
В
—
гранулярна
ендоплазматична
сітка.
в
що вони розташовані в цих оболонках двома шарами. При цьому ліпідні молекули повернуті одна до одної гідрофобними ланцюгами жирних кислот. На зовнішній та внутрішній поверхнях мембрани розміщені полярні гідрофільні головки ліпідних молекул. Основою мембрани є подвійний шар ліпідних молекул. Зовнішня та внутрішня поверхні біліпідного шару утворені гідрофільними частинами і вкриті суцільними шарами білка.
Ще в 50-х роках xx ст. цю модель досліджували електронно-мікроскопічним способом. у подальшому Робертсон запропонував гіпотезу про єдину унітарну біологічну мембрану. Однак численні морфологічні і функціональні дослідження клітинних мембран не підтвердили цієї гіпотези. Найімовірнішою вважається рідинно-мозаїчна модель. Тут в основі мембрани той самий біліпідний шар. Білки, які входять до складу мембрани, не утво-
рюють суцільного шару. їх можна розподілити на три групи: периферійні, інтегральні та напівінтегральні.
Периферійні білки розташовані на зовнішній та внутрішній поверхнях біліпідного шару і з'єднуються з його полярними головками електростатичними силами. Вони пов'язують мембрану з над- і субмембранними структурами поверхневого апарату клітини.
Інтегральні та напівінтегральні білки мають глобулярну структуру. Вони пов'язані з ліпідами гідрофільно-гідрофобними взаємодіями. Інтегральні білки пронизують усю товщу біліпідного шару. Напівінтегральні білки занурені в мембрану лише частково: вони виступають на її зовнішній або внутрішній поверхні.
Глобули можуть зміщуватися латерально і вертикально, тобто мембрана є лабільною, динамічною, має термодинамічну стійкість. Для підтримки не потрібні затрати енергії.
Нові морфобіохімічні дослідження підтвердили можливість такої будови мембран.
Серед ліпідних мембран виділяють дві основні групи: структурні та регуляторні. Перші є основою мембрани, а другі беруть участь у функціях глобулярних білків.
Мембранні білки здійснюють рецепторні функції. Білки — активні та пасивні переносники сполук.
До внутрішньої поверхні клітинної мембрани прилягає зовнішній шар цитоплазми — субмембранна система гіалоплазми.
у клітині є два види транспортних процесів:
пасивний транспорт, або полегшена дифузія, за градієнтом концентрації;
активний транспорт з пересуванням речовин проти градієнта концентрації.
Перенесення речовин здійснюють дві групи мембранних транспортних білків: білки-канали та білки-переносники.
Білки-канали переносять йони значно швидше, ніж білки-переносники.
Мембранний транспорт різних оформлених частинок у клітині відбувається шляхом ендоцитозу (фаго- і піноцитозу) та екзо-цитозу.
Екзоцитоз — це вихід речовин з клітини у вигляді гранул секрету або вакуоль з клітинною рідиною.
У рослинній і бактеріальній клітинах із зовнішнього боку мембрани є товста стінка, яка здійснює функцію опори. Ця клітинна стінка у рослин містить целюлозу, а в бактерій — муреїн.
Головна функція клітинної мембрани — забезпечити надходження в клітину речовин і зберегти постійність її складу (клітинний гомеостаз). Вона є захисним молекулярним бар'єром, тобто здійснює контроль за вибірною проникністю в клітину та з клітини певних речовин, забезпечує здатність до фаго- та піноци-тозу, а також зв'язок між клітинами в тканинах багатоклітинного організму.
Цитоплазма становить основну масу клітини. У світловому мікроскопі цитоплазма представлена гомогенною, безбарвною, прозорою слизистою рідиною. В убитих фіксованих клітинах цитоплазма має ниткоподібну зернисту або пінисту структуру.
Електронномікроскопічними дослідженнями встановлено, що складові живої клітини знаходяться в стані безперервного руху і взаємодії, унаслідок чого її внутрішня структура часто змінюється.
У цитоплазмі розрізняють гіалоплазму — цитоплазматичний матрикс, гомогенне середовище, де розміщені клітинні органе-ли, яким властивий єдиний життєвий саморегулювальний процес. З гіалоплазмою зв'язані колоїдні властивості цитоплазми, її в'язкість, скоротність, внутрішній рух. Гіалоплазма складається з білкових ниток та зернистих утворень, багатих на рибонуклеїнові кислоти. До її складу входять також різноманітні ферменти. Різні ділянки гіалоплазми мають неоднакову макромолекулярну структуру.
Для гіалоплазми характерна також наявність цитоскелета, який складається зі спаяних між собою міцел.
Ядро — центральний інформаційно-генетичний апарат клітин. Воно являє собою колоїдні системи, до складу яких входять вода, органічні сполуки, особливо нуклеїнові кислоти, білки та мінеральні солі. Як правило, у клітині ядро одне, проте деякі клітини (наприклад, кісткового мозку та ін.) можуть мати двоє або більше ядер.
Ядро — обов'язковий компонент усіх клітин, за винятком лише деяких високоспеціалізованих, які втратили здатність до поділу і мають нетривалий період життя (наприклад, зрілі еритроцити; незрілі — мають ядра). Ядро відділене від цитоплазми ядерною мембраною (оболонкою). Мембрана двошарова, у ній є пори, крізь які вміст ядра сполучається з цитоплазмою, але ніколи не змішується. Між цитоплазмою і ядром обмін речовин відбувається крізь пори.
Розміри і форма ядра залежать від форми, розмірів і функції клітини. Найчастіше ядро округле, паличкоподібне. Деякі клітини мають двоє ядер і більше (наприклад, клітини печінки, м'язів людини).
До складу ядра входять ДНК, РНК, білки. У ядрі зосереджено 99 % ДНК клітини. В ядрі ДНК разом з білками утворює комплекси дезоксирибонуклеопротеїдів (ДНП).
Зовнішня мембрана ядра переходить в ендоплазматичну сітку (ЕІІС). Ядерна оболонка виконує захисну функцію і регулює обмін речовин.
Ядро заповнене ядерною речовиною (каріоплазмою). До її складу входять різні види РНК, білки, нуклеотиди, амінокислоти, ферменти. Крім того, у ній містяться одне або два ядерця (в ядерці, у свою чергу, виявлено РНК та білки; воно виконує функцію синтезу субодиниць рибосом), а також глибки і гранули хроматину. Хроматин (забарвлений) — це комплекс ядерної ДНК, РНК зі структурними білками — гістонами та негістоновими білками. Із хроматину в період поділу клітини формуються інтенсивно забарвлені структури — хромосоми (мал. 4).
Основні функції ядра:
зберігає, відтворює і передає спадкову інформацію;
реалізує спадкову інформацію;
керує всіма процесами життєдіяльності клітини;
бере участь у розмноженні;
регулює обмін речовин у клітині.
К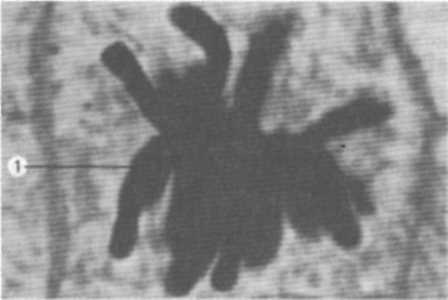 ожний
вид
рослин
і
тварин
має
певну
і
постійну
кількість
хромосом.
Для
хромосом
характерні
правила
парності,
індивідуальності,
безперервності.
ожний
вид
рослин
і
тварин
має
певну
і
постійну
кількість
хромосом.
Для
хромосом
характерні
правила
парності,
індивідуальності,
безперервності.
Соматичні клітини тіла мають повний, подвійний (диплоїдний) набір хромосом (2п), а статеві – одинарний (гаплоїдний) набір
Мал. 4. Метафазні хромосоми (1) у рослинній клітині
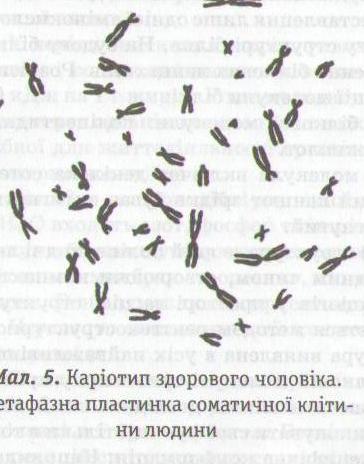
хромосом (n). Хромосомні набори соматичних клітин чоловічої та жіночої статей одного виду відрізняються між собою за однією парою хромосом. Ця пара називається статевими хромосомами. Решта пар хромосом відносно однакової морфології в обох статей називається аутосомами. Так, у людини 22 пари аутосом та 23-я пара статевих, або гетерохромосом.
Диплоїдний набір хромосом, виділений із клітин (у метафазі мітозу), який характеризується формою, розміром і кількістю, називається каріотипом (мал. 5), а попарне розташування хромосом клітини в порядку зменшення їх розмірів — ідеограмою.
Основа хромосоми — довга молекула ДНК, завжди в комплексі з білками, частіше — з гістонами, утворює нуклеопротеїди. Поряд з ними є й інші (кислі) білки, а також РНК. У молекулах ДНК записана спадкова інформація клітини.
Ядерця — утворення в інтерфазних ядрах більшої частини рослинних та тваринних клітин.
Кількість ядерець дорівнює від 2—3 до кількох десятків. Типова форма — округла, однак можуть бути різні варіанти. Згідно з даними електронної мікроскопії, ядерця не мають обмежувальної мембрани і є сіткою ниток — нуклеонем. Останні складаються з субмікроскопічних гранул, вміщуючи рибосомну РНК.
У період мітозу ядерця зникають, а після його закінчення виникають знову. Вони утворюються на спеціальних ділянках хромосом, які звуться ядерцевими організаторами. Нитки ДНК ядер-цевого організатора наскрізь пронизують ядерця і є його головним функціональним компонентом. На них синтезується рибосомна РНК, яка тут-таки, в ядерцях, включається до складу утворених рибосом. Крім того, синтезуються основні білки.
Морфологія клітини
Органели загального і спеціального призначення
Основна частина цитоплазми, в якій розміщені органели та включення, — це гіалоплазма.
Органели це постійні високодиференційовані внутрішньоклітинні утворення, які виконують певні функції. Органели поділяють на групи загального (ендоплазматична сітка, рибосоми, лізосоми, мітохондрії, пластинчастий комплекс, внутрішньоклітинний апарат, центросоми, мікротрубочки та мікротільця, пластиди) і спеціального (війки та джгутики, міофібрили, мікровор-синки, кореневі волоски) призначення.
Органели малих розмірів відкрито за допомогою електронного мікроскопа. Це система внутрішньоклітинних мембран, які пронизують клітинний вміст в усіх напрямках. Мембрани у середньому завдовжки 5—10 нм.
Ендоплазматична сітка є в цитоплазмі всіх клітин тварин і рослин, за винятком яйцеклітин деяких груп тварин, в яких ця органела недостатньо розвинена або не виявляється, а також в одноклітинних організмів. Вона буває двох типів: гранулярна (шорстка) і агранулярна (гладка). На мембранах каналів шорсткої (гранулярної) ендоплазматичної сітки, або ергастоплазми, розміщуються рибосоми. Канали гранулярної сітки можуть бути зв'язані із зовнішньою плазматичною мембраною клітини, а також з оболонкою ядра і найбільш розвинуті в клітинах спеціалізованих тканин (підшлункової та слинних залоз, печінки та ін,). Основна функція гранулярної ендоплазматичної сітки — участь у синтезі білка з подальшим проникненням його в різні ділянки клітини. Вона утворює систему мембран і каналів, зв'язаних з основним шаром цитоплазматичної мембрани. Діаметр каналів досягає 10—20 нм. Добре розвинута гранулярна сітка в клітинах, які виробляють речовини ліпідного характеру. На її мембранах відбувається синтез жирів, вуглеводів і гормонів. Роль ендоплазматичної сітки в житті клітини дуже важлива, тому що вона означає циркуляційну течію цитоплазми і бере участь в обмінних процесах унаслідок наявності на її мембранах численних ферментних систем.
Розмір, форма, взаєморозміщення структурних компонентів ендоплазматичної сітки в кожного виду клітин специфічні. Однак названі параметри можуть досить сильно варіювати залежно від функціонального стану клітини. Ендоплазматична сітка досить швидко реагує на ушкодження, що діються в клітині.
Рибосоми (рибо — від назви РНК, яота — тіло) виявлені за допомогою електронної мікроскопії в клітинах усіх організмів, починаючи від бактерій і закінчуючи ссавцями. Це маленькі елек-троннощільні сферичні утворення завбільшки 8—35 нм, прилеглі до зовнішньої поверхні мембран ендоплазматичної сітки. Аналогічні гранули можна спостерігати вільно лежачими в основному цитоплазматичному матриксі і прилеглими до зовнішнього шару оболонки ядра. Цих гранул немає ні на мітохондріях, ні на плазматичній мембрані. У великій кількості розміщені в клітинах, які активно синтезують білки.
Складовою частиною рибосом є білки, РНК. За допомогою ультрацентрифугування встановлено, що рибосоми можуть розпадатися на дві нерівні субодиниці. У меншу субодиницю молекулами РНК транспортуються амінокислоти, а в більшій локалізується поліпептидний ланцюжок, що зростає.
Рибосоми можуть скупчуватися у вигляді розеток або кружалець. Виявлені численні групи, які складаються, наприклад, з 5—7 рибосом, а інколи чітко видно найтоншу нитку іРНК, яка об'єднує рибосомальні одиниці. Таке об'єднання рибосом у групи утворює полірибосоми (полісоми). Синтез білка (основна функція цих органел) відбувається не на ізольованих рибосомах, а на їхніх комплексах.
Лізосоми (від грец. lysis — розчинення, soma — тіло) — органели, виявлені майже в усіх тваринних клітинах, включаючи найпростіших. Найбільш визначеним типом первинних лізосом є гранули сегментноядерних нейтрофілів. Лізосоми — це округлі електромагнітні утворення діаметром 0,2—0,8 мкм.
Лізосоми багаті на ферменти (їх описано понад 400 видів). Унаслідок дії ферментів лізосом складні органічні сполуки розщеплюються на прості. Наприклад, білки розпадаються до амінокислот.
Первинні лізосоми походять від внутрішнього сітчастого апарату, який є важливим джерелом утворення пухирців у клітині. Лізосоми, як фагосоми або піносоми, також являють собою мікро-пухирці, але, на відміну від первинних не містять у собі ферменти, необхідні для внутрішньоклітинного травлення.
Вторинні лізосоми виникають унаслідок злиття первинних лізосом із фагосомами. У результаті формуються травні вакуолі, в яких здійснюється внутрішньоклітинне травлення.
Після закінчення процесів травлення і всмоктування поживних речовин травні вакуолі перетворюються на залишкові тільця, їхній вміст — залишки поглинутих і неутилізованих речовин. Утворені залишкові тільця підходять до цитоплазматичної мембрани і виводяться назовні. Виникає процес, прямо протилежний фагоцитозу, — екзоцитоз. Лізосоми можуть виникати внаслідок відторгнення мембрани від уражених ділянок клітини. Ці структури близькі до травних вакуоль, але функція у них інша. Вона спрямована на захист клітини від її уражених елементів. При цьому перетравлюються непотрібні ділянки клітини. Тоді лізосоми відокремлюють ділянки, які підлягають перетравлюванню, і виділяють туди свої ферменти. Описано випадки самоперетравлювання клітини внаслідок дії власних лізосом.
Смерть клітин і дегенерація їхніх субстанцій — неминуче явище під час розмноження та утворення нової клітини. У цьому зв'язку лізосоми також беруть участь у процесах морфологічної еволюції метаморфозу, дегенерації і регенерації тканин. Велика роль лізосом і в клітинній патології. Вони є одночасно і компонентом імунологічних процесів, які відбуваються в організмі.
Мітохондрії виявлені за допомогою мікроскопа в усіх клітинах (за винятком еритроцитів) рослин, тварин і бактерій. Це ліпопро-теїнові утворення різної величини (0,5—0,6 мкм), дуже різноманітні за своєю будовою. На фіксованих препаратах вони помітні як нитки або зерна. Від цього й походить їхня назва.
Зовнішня оболонка мітохондрій складається з двох твердих щільних мембран завтовшки 6 нм, розміщених на відстані 6— 8 нм одна від одної. Від внутрішньої мембрани оболонки відходять внутрішні складки-кристи, гребені. Останні в середині мітохондрій утворюють поперечні або навкісні перегородки. Мінливість мітохондрій стосується не тільки форми, а й внутрішньої структури. Велика кількість поперечних гребенів виявляється в міто-хондріях м'язових клітин. В окремих видів найпростіших гребені трубчастої форми. Утворена цими структурами порожнина заповнена основною речовиною — матриксом. Поверхня внутрішньої мембрани і крист має елементарні частинки, розміщені на ніжках, величиною 8—10 нм. Вважають, що вони беруть участь у перенесенні електронів. Кількість мітохондрій залежить також від фізіологічного стану клітини. У період голодування кількість крист мітохондрій клітин печінки зменшується, мітохондрії набрякають, а їхній матрикс світлішає.
Мітохондрії здатні до розмноження. Вони мають у своєму складі мітохондріальну ДНК, яка несе спадкову інформацію про будову і функцію цих органел. У зиготу мітохондрії потрапляють із цитоплазмою яйцеклітини.
У мітохондріях синтезується АТФ, яка є основним джерелом енергії в клітині. Органічні речовини (продукти розпаду вуглеводів, білків, жирів) надходять у мітохондрії з цитоплазми і проходять там кілька окисних реакцій, кінцевим результатом яких є утворення вуглекислого газу і води. Процес (цикл Кребса, або трикарбонових кислот, або фосфорилювання в дихальному ланцюгу) здійснюється за участю груп ферментів, а також особливих речовин — переносників електронів. Цей процес є фосфорилюван-ням АДФ і здійснюється з деякими окисно-відновними реакціями аеробної фази метаболізму. Унаслідок окиснення органічних речовин у мітохондріях вивільнюється велика кількість енергії, яка перетворюється на теплову, але близько 50 % її акумулюється і зберігається у вигляді хімічної енергії. Це зумовлено тим, що паралельно циклу Кребса в мітохондріях відбуваються процеси, результатом яких є створення АТФ, АДФ та фосфату.
АТФ бере участь в усіх внутрішньоклітинних процесах, які здійснюються з використанням енергії. При цьому вона розпадається на АДФ і фосфат, розривається макроергічний зв'язок, тоді як акумульована в ньому енергія використовується клітиною. Отже, АТФ є акумулятором, зарядження якого відбувається в мітохондріях, а розрядження — в інших ділянках клітини в період метаболічного процесу. Утворення АТФ у мітохондріях унаслідок використання енергії описаних реакцій називають окисним фос-форилюванням.
Поряд зі своєю основною енергетичною функцією мітохондрії виконують і інші. Наприклад, вони накопичують деякі речовини, які надходять до неї в підвищених кількостях. У них синтезуються жирні кислоти. Мітохондрії регулюють концентрацію солей у цитоплазмі. Крім того, їм властива і генетична функція, яка зумовлена наявністю в них молекул ДНК, котрі є носіями спадкової інформації. У мітохондріях синтезується багато власних білків. З експериментальних даних відомо, що явище цитоплазматичної спадковості значною мірою пов'язане з генетичною діяльністю мітохондрій.
Мітохондрії беруть участь у секреції, накопиченні жиру, глікогену, хлорофілу, а також у змінах, зумовлених впливом їжі (в печінці), у реабсорбції (в нирці) або в загальному метаболізмі. Вони зазнають змін у процесі розвитку і в патологічних чи перед-патологічних умовах.
Комплекс Гольджі — внутрішній пластинчастий апарат — наявний в усіх клітинах рослинних і тваринних організмів. Він складається зі сплющених цистерн (мішечків, розміщенних паралельно у вигляді штабелів), наповнених системами з подвійними стінками (мембранами). Цистерни розміщені комплексно по 4—8 пар, просвіток однієї цистерни сягає 15—ЗО нм. Комплекс Гольджі також містить різної величини вакуолі і маленькі пухирці. Великі вакуолі, створені внаслідок розширення деяких цистерн, легко відрізнити від дрібних пухирців. Великі вакуолі розташовані на периферії комплексу Гольджі, на його зовнішній, дистальній поверхні і здатні гіпертрофуватися. Вміст великих ва-куолей складається з води, на підставі всього цього деякі цитологи вважають, що вони беруть участь у водному обміні. Дрібні пухирці обмежені тонкою мембраною. Вони згруповані по сусідству зі сплющеними цистернами, з їх продовженням, утвореним унаслідок відбруньковування.
У хімічному відношенні значне місце у внутрішньому сітчастому апараті належить ліпідам, особливо фосфоліпідам, а також білкам. Існують кислі і лужні фосфатази: нуклеодифосфатаза, ті-амінпрофосфотаза та інші ферменти.
Функція комплексу Гольджі пов'язана з основними процесами обміну, які стосуються її секреторної активності. Є дані про зв'язок комплексу Гольджі із загальним обміном. Так, при насиченому клітинному метаболізмі комплекс Гольджі досить сильно розвинутий і добре помітний. Це явище характерне не тільки для нормальних клітин, а й для клітин у період генезису, перед патологічним підвищенням метаболізму. Під впливом окремих фармакологічних або гормональних препаратів нарівні з підвищенням загального обміну клітини виявляються ознаки, які характеризу-ютьстимуляціюфункції комплексу Гольджі. Комплекс Гольджі — місце синтезу різних складних вуглеводів, які виконують різні фізіологічні функції. Ці вуглеводи утворюють слиз, що захищає кишки від впливу бактеріальної флори. Вони входять до складу продуктів, які виробляють підшлункова залоза, прищитоподіб-ні та слинні залози, до складу зубної емалі та основної речовини хряща.
Повідомлення про фізіологічний зв'язок комплексу Гольджі з ядром або лізосомами потребують підтверджень.
Клітинний центр (центросому) виявлено в усіх клітинах, за винятком нервових, яйцеклітин та клітин вищих рослин. Орга-нела складається з одного або двох твердих тілець — центріолей, які добре сприймають барвники, та навколишньої світлої зони — центросфери. Згідно з даними електронномікроскопічних досліджень, центріоль має вигляд циліндра діаметром до 160 нм, стінки якого складаються з 9 трубочок або паличок завтовшки 300— 500 нм; центріоль має відносно тверду субстанцію. Зазначена кількість паличок постійна в усіх клітинах усіх видів організмів.
З трубочками пов'язані округлі тільця — сателіти, або перицен-тріолярні тільця, які не завжди виявляються. Центріолі беруть участь у мітозі і нагадують структури, розміщені біля основи війок та джгутиків, які є в клітинах. Відомо, що центріолі містять у собі фермент, під дією якого відбувається розпад аденозинтри-фосфорної (АТФ) кислоти — основного акумулятора енергії клітини. В інтерфазі центріолі завжди мають стандартний вигляд, позбавлені функціонального значення і наявні лише як резерв.
Мікротрубочки та мікротільця. Мікротрубочки виявлені в клітинах усіх тваринних та рослинних організмів. Це циліндричні, ниткоподібні утворення завдовжки 20—ЗО мнм, діаметром близько 23 нм. Оболонка мікротрубочок тришарова, завтовшки близько 5 нм. Розташовуючись у цитоплазмі, вони надають клітині відповідної форми і забезпечують стійкість та стабільність локалізації органел, а також беруть участь у транспортуванні води, йонів та деяких молекул.
Мікротільця — дрібні овальні утворення розміром близько 0,5 мнм з одношаровою мембраною і стрижнеподібною кристалічною структурою. Виявлено мікротільця в одноклітинних, багатоклітинних організмах, а також у клітинах дріжджів та в людини.
Ці утворення, як і інші компоненти клітин, містять у собі ферменти. Універсальним ферментом для всіх клітин є каталаза, але вони містять у собі також уротоксидазу, оксидази амінокислот.
Пластиди виявлені тільки в клітинах рослинних організмах. У світловому мікроскопі пластиди виявляються у вигляді паличок, лусочок, зерен завбільшки 1 —12 мкм, а при електронномікро-скопічному дослідженні установлено, що, наприклад, хлоропласт має подвійну зовнішню мембрану. Внутрішня структура також представлена мембранами, між якими розташовані грані у вигляді зерен, утворених мішечками з подвійних мембран, які щільно прилягають один до одного. Виникають пластиди пропластид — округлих тілець розміром 1—2 мкм, які мають подвійну мембрану. Кількість пластид збільшується шляхом їх поділу.
Розрізняють такі види пластид: зелені (хлоропласти), жовті, оранжеві, червоні (хромопласти) та безбарвні (лейкопласти). Пластиди можуть перетворюватися з одного виду на інший: лейкопласти — на хлоропласти, а потім — на хромопласти. Найбільший інтерес становлять хлоропласти, оскільки вони забезпечують явище фотосинтезу, за допомогою якого синтезуються АТФ та глюкоза.
Спільність у структурі хлоропластів і мітохондрій зумовлює і спільність їхніх функцій. Якщо в мітохондріях відбувається трансформація енергії, яка збільшується в процесі дисиміляції, то в хлоропластах здійснюється фотосинтез. У результаті сонячна енергія трансформується в хімічну.
Хромопласти містять у собі каротиноїди, білки, ліпіди, вітаміни Е, К, Б, а також до 4 % РНК. Отже, у них здійснюються різні синтетичні процеси і крім глюкози утворюються білки, ліпіди та деякі вітаміни.
Органели спеціального призначення — це кореневі волоски, війки, джгутики. Наприклад, війки миготливого епітелію, джгутики сперматозоонів та деяких вільноживучих і паразитичних найпростіших класу Р1а£е11аІа, а також бактерій — джгутиконосців.
Клітинні включення. Нарівні з органелами в протоплазмі клітин можна знайти включення трофічного, секреторного та спеціального призначення. Вони являють собою продукти внутрішньоклітинного обміну або запасні живильні речовини. Залежно від хімічної природи розрізняють білкові, жирові та вуглеводні включення.
Хімічний склад клітини
У клітині виявлено понад 70 із 110 елементів періодичної системи Д.І. Менделєєва. Клітина живої істоти складається з тих самих елементів, що й об'єкти неживої природи. Це вказує на спільність живої та неживої природи. Елементи періодичної системи, які беруть участь у метаболізмі і мають виразну біологічну активність, називаються біогенними.
До хімічного складу цитоплазми рослинних і тваринних клітин входять близько 70 % кисню, 18 % вуглецю, 10 % водню, що в сумі становить 98 % загальної живої маси клітини.
На частку азоту, кальцію, кремнію, натрію в клітинах організму припадають десяті частки відсотка, а фосфору, магнію, сірки, хлору, алюмінію, заліза — соті частки відсотка. Названі елементи періодичної системи разом із киснем, вуглецем і воднем становлять 99,99 % маси клітини.
Марганець, бор, мідь, цинк, фтор, барій, нікель, літій, йод, кобальт, хром становлять тисячні, десятитисячні, стотисячні частки відсотка загальної маси клітини; вони називаються мікроелементами. Мільйонні частки відсотка в клітині становлять ртуть, золото, радій тощо — це ультрамікроелементи. Вони мають велику біологічну активність, беручи участь в основних метаболічних процесах. Мікроелементи є складовою частиною мета-лоферментів, гормонів, вітамінів та інших біополімерів. Вони є також стимуляторами імуногенезу, виявляючи при цьому виражену активність, що впливає на систему неспецифічного імунітету, процеси антитілоутворення. Завдяки цьому мікроелементи використовують у профілактиці й терапії певних захворювань (хвороб дитячого віку, атеросклерозу, анемії, ендемічного зоба та ін.). Наприклад, додаткове введення експериментальним тваринам мікроелементів міді, марганцю, кобальту посилює антитіла утворювальної системи лімфоїдної тканини, що змінює реактивність організму щодо реакційного агента. Отже, значення того чи іншого елемента полягає не тільки в його кількості.
Як і мікроелементи, ультрамікроелементи, виявлені в живих істотах, необхідні для життя. Відсоткове співвідношення макро-та ультрамікроелементів різних організмів варіабельне, що залежить від виду організму.
Типи хімічних зв'язків та їх значення. Ионний, або електростатичний, зв'язок виникає в результаті притягання частинок з позитивними і негативними зарядами. Наприклад, Na+, Cl, NaCl. Йонний зв'язок в організмах притаманний в основному неорганічним сполукам.
Ковалентний зв'язок утворюється внаслідок узагальнення електронної пари двох різних атомів, які відзначаються міцністю, і потребує для свого утворення енергії. Виникнення і розрив цього зв'язку здійснюється в перебігу ферментативних реакцій. Прикладом ковалентного зв'язку може бути пептидний:
Н2N-СН2-СООН + Н2N—СН2—СООН =
= Н2NCН2—СОNH—СН2—СООН + Н2О.
Водневий зв'язок — це, за суттю, також йонний зв'язок, або електростатичний, але більш слабкий і виникає між двома сильно негативними атомами. Наприклад, при асоціації спирту або при міжмолекулярній взаємодії спиртів з амінами:
CH2-O –H....N H2C6H5
Ці типи хімічних зв'язків зумовлюють виникнення нових молекул з молекул, здатних реагувати. Унаслідок цього в клітині здійснюються найскладніші процеси метаболізму. Усі виявлені в живих істотах хімічні елементи є складовою частиною хімічних сполук. Одні з них неорганічні, інші — органічні.
Із неорганічних речовин до складу живих істот входять вода, мінеральні солі і деякі неорганічні кислоти. Вода має виключно важливе значення в життєвих процесах. Речовини цитоплазми утворюють з водою справжні або колоїдні розчини. У цих водних розчинах проходять усі хімічні та фізико-хімічні реакції. Вода — це не просто розчин, а активний учасник багатьох процесів метаболізму клітини. За кількісним вмістом у цитоплазмі (у середньому 70—80 %) вона стоїть на першому місці. Тіло дорослої людини містить у собі 60 % води, а людський ембріон — у межах 97 %.
У цитоплазмі вода перебуває у вільному і зв'язаному станах як складова частина різних сполук. Основна маса (близько 95 %) води в клітинах знаходиться у вільному стані і відіграє роль розчинника хімічних речовин і середовища, де відбуваються важливі хімічні реакції (ферментативні, окисні, виведення різних речовин з клітини та ін.). У зв'язаному стані перебуває 4—5 % всієї маси води. Зв'язана вода складається з трьох молекул, які водневими та іншими зв'язками неміцно з'єднані з білками. Разом з тим зв'язана вода міцно утримується колоїдними часточками, не розчиняє солей і замерзає за температури -40 С.
Найважливіші фізичні властивості води — теплоємність, теплопровідність, великий обсяг тепла пароутворення. Завдяки цим властивостям вода добре охороняє протоплазму від різних змін температури, забезпечує рівномірний розподіл тепла і захищає організм від перегрівання. Вона також є мастилом у процесі тертя внутрішніх органів.
Взаємозв'язок інтенсивності обміну і кількості води вказує на її велике біологічне значення. Так, води в сірій мозковій речовині міститься в межах 86 %, у нирках — 70 %, тоді як у кістках — 22 %, а в емалі зубів — близько 10 %. Зменшення інтенсивності процесів метаболізму веде за собою і зменшення кількості води. Без води хід життєвих процесів неможливий. З цієї причини кількість води, яка зменшується в результаті випаровування, обов'язково має поповнюватися, інакше гине життя, жива істота. Разом з тим деякі найпростіші організми тривалий час зберігають життєдіяльність у висушеному стані (явище анабіозу).
Мінеральні солі відіграють важливу роль у життєдіяльності організму. На них припадає 2—8 % сухої речовини клітини.
Солі натрію, калію, кальцію, магнію, хлорної, сульфатної, фосфорної та інших кислот, незважаючи на порівняно невеликий відсотковий вміст їх у клітині, впливають на рівень осмотичного тиску, забезпечують сталість кислотності та лужності цитоплазми. Виступаючи як електроліти, розчини солей впливають на колоїдний стан білків. Нормальний хід життєдіяльності організму потребує абсолютного і повного співвідношення різних солей у цитоплазмі; зрушення в бік зменшення або збільшення веде до загибелі організму.
Органічні речовини. Вуглеводи. Складовими частинками молекул вуглеводів є С, Н, О. Вуглеводи поділяють на три основних класи:
Моносахариди (глюкоза, або виноградний цукор, СбН1206) — первинний енергетичний матеріал клітини.
Дисахариди (сахароза С12Н22Оп, мальтоза у рослин, лактоза у тварин). При гідролізі кбжна молекула дисахариду розщеплюється на дві молекули моносахариду.
Полісахариди — утворюються в результаті сполучення значної кількості ділянок молекул моносахаридів. Загальна формула полісахаридів — (С6Н10О5)п, де п - кількість молекул моносахаридів. У період гідролізу полісахариди розпадаються на моносахариди. До полісахаридів належать крохмаль, глікоген (тваринний крохмаль), целюлоза, клітковина.
Вуглеводи є складовою частиною цитоплазми. Вони відкладаються в цитоплазмі у вільному стані як джерело енергії організму. При окисненні 1 г вуглеводів утворюється 27,6 кДж теплової енергії. Вуглеводи входять до складу мембранних структур клітини, беруть участь у двох бар'єрних функціях. Утворюючи складні комплекси з білками, входять до багатьох видів сполучної тканини, хряща, кісткової тканини, а також до складу оболонок яйцеклітини, тканини тестикули, склистого тіла ока та ін. Моносахариди, дисахариди, полісахариди, вступаючи в сполуки з іншими речовинами клітини, зумовлюють її колоїдні та осмотичні властивості.
Ліпіди (жири та жироподібні розчини — ліпоїди). Жири, як і вуглеводи, безазотисті органічні речовини. До їх складу також входять С, Н, О, але в інших співвідношеннях. Жири — це сполуки гліцерину з жирними кислотами (олеїновою, стеариновою та ін.). У воді жири не розчиняються, а утворюють емульсії. Продукти гідролізу жиру — гліцерин і жирні кислоти. Організми здатні переводити вуглеводи в жири і накопичувати енергетичний запас.
У період окиснення жирів, унаслідок утворення вуглекислого газу і води, вивільнюється велика кількість теплової енергії (40 кДж на 1 г жиру). У цьому відношенні жири посідають перше місце серед органічних сполук і є важливим джерелом енергії, потрібної для життєдіяльності організму; виконують також функцію термоізоляції та ін.
Ліпоїди — жироподібні речовини, до складу молекул яких крім С, Н, О входять азот, фосфор та інші елементи. Ліпоїди у вигляді сполук з білками утворюють ліпопротеїди, які входять до складу клітинних мембран та органел, що мають мембранний принцип будови. Вони зумовлюють проникність клітинних мембран.
Нервова тканина, яєчний жовток, сперма — також багаті на ліпопротеїди.
Білки називають також протеїнами (від грец. ргоіе — перший). Основна роль в організмі належить білкам, з якими завжди пов'язані всі прояви життя. Безліч хімічних реакцій метаболізму, реакцій синтезу одних сполук і розпаду інших здійснюється в організмі внаслідок каталітичної функції білків.
До складу білків входять атоми С, Н, О, N та незначна кількість 8, Р, Ге і деяких інших елементів. Складаються білки з амінокислот, які характеризуються наявністю в їхній молекулі аміногрупи — 1*Ш2 і карбоксильних груп — СООН, унаслідок чого амінокислоти мають властивості і лугів, і кислот (амфотерність). Завдяки цим особливостям амінокислоти легко з'єднуються одна з іншою через СООН і МН2. Утворений при цьому зв'язок СО—N11 називається пептидним. Поєднання двох амінокислот — це дипептид, трьох — це трипептид. Пептид, який складається з невеликої кількості амінокислот, називається олігоиептидом, а з багатьох — поліпептидом.
До складу білкових молекул входить 20 різних амінокислот, які є найнеобхіднішими для організму. Наприклад: тирозин, аланін, лейцин, аргінін, лізин та ін. Загальна кількість їх у молекулі різних білків неоднакова (до 300 і більше). У різних білків також неоднаковий якісний склад амінокислот.
За хімічною структурою білки — біологічні полімери, в яких амінокислоти йдуть послідовно у вигляді намистинок на нитці. Вільні амінокислоти клітини утворюються внаслідок розщеплення білків або надходять з міжклітинної рідини і становлять амінокислотний фонд, який є будівельним матеріалом для синтезу нових білків. Заміна або переставлення лише однієї амінокислоти призводить до значних змін у структурі білка. На будову білків впливає і просторове розміщення білкових ланцюжків. Розрізняють декілька рівнів організації молекули білків:
Первинна структура білкової молекули: поліпептидний ланцюг побудований із амінокислот.
Вторинна структура: молекула включає декілька сотень амінокислот, і поліпептидний ланцюг зрідка буває витягнутим повністю. Як правило, він зігнутий.
Третинна (глобулярна) структура, в якій поліпептидні ланцюги скорочуються відповідним чином, створюючи компактну структуру. Розміщення ланцюгів у просторі за цієї структури білка дуже складне і вивчається методом рентгеноструктурного аналізу. Глобулярна структура виявлена в усіх найважливіших білках. Вона зумовлює в білковій молекулі тривимірну просторову конформацію. Експериментальними дослідженнями встановлено, що білок здатний виконувати свої функції тільки в тому випадку, коли збережена специфічна конформація. Наприклад, якщо в молекулі білка розірвати зв'язок, який утворює його третинну структуру, то ферментативні властивості білка повністю втрачаються. У визначенні конформації білкової молекули велике значення має співвідношення гідрофільних і гідрофобних амінокислот. Гідрофільні сполуки характеризуються наявністю водневого зв'язку в молекулі води. Ці сполуки легко вступають у реакції з водою. Гідрофобні ж сполуки, навпаки, погано вступають у реакції з водою через наявність вуглеводистих груп СНЯ, СдНб та ін. Отже, конформація білкової молекули, її вторинна і третинна структури визначаються її первинною структурою.
Четвертинна структура: два абобільше ланцюгів, які можуть бути однаковими або різними, але в обох випадках вони пов'язані слабкими зв'язками (нековалентними). Наприклад, молекула гемоглобіну складається з чотирьох поліпептидних субодиниць (двох Ь- і В-ланцюгів), поділ і асоціація яких можуть відбуватися спонтанно. Багато ферментів та інших білків з молекулярною масою понад 50 000 характеризуються четвертинною структурою. Існують складні білки, до молекул яких входять прості білки та небілкова частина, — це протеїди. До них належать нуклеопро-теїди (білки, зв'язані з нуклеїновими кислотами), ліпопротеїди (наприклад, ліпопротеїди крові), хромопротеїди (гемоглобін).
 гемоціанин
і
цитогарми),
в
яких
білок
зв'язаний
з
ферментом,
та
ін.
Особлива
роль
належить
нуклеопро-
теїдам,
які
є
в
усіх
клітинах.
гемоціанин
і
цитогарми),
в
яких
білок
зв'язаний
з
ферментом,
та
ін.
Особлива
роль
належить
нуклеопро-
теїдам,
які
є
в
усіх
клітинах.
Білки в клітині виконують різні функції (структурну, сигнальну, рушійну, ферментативну та ін.).
Для кожного виду організмів характерний
свій видовий специфічний білок. Існує не тільки видова, а й індивідуальна
специфічність білків.
Навіть у різних тканинах одного й того
самого організму
виявляється тканинна специфічність білків. Виявлення специфічності білків здійснюється за допомогою біологічних (імунологічних) методів. Уведення в організм стороннього білка (антигену) стимулює утворення специфічних білків-антитіл, які вступають у взаємодію саме з цим білком. Звідси можна зробити висновок, що утворення в організмі антитіл у результаті введення стороннього білка — явище біологічне і корисне. Разом з тим воно є суттєвою перешкодою в трансплантації органів і тканин.
Нуклеїнові кислоти (від лат. писіеиз — ядро) — хімічні речовини, які мають важливе біологічне значення, відіграють провідну роль у біосинтезі білка, спадковості та інших життєво важливих процесах. Нуклеїнові кислоти — високоспецифічні полімери, їх макромолекули — це довгий одинарний або подвійний ланцюг, утворений безліччю ділянок (мономерів), які в нуклеїнових кислотах називаються нуклеотидами.
Відомо дві групи нуклеїнових кислот: дезоксирибонуклеїнові (ДНК) і рибонуклеїнові (РНК). ДНК розміщуються головним чином у ядрі клітини і є основною речовиною хромосоми; РНК — у ядрі, цитоплазмі, частково в хромосомах. ДНК складається з двох спірально закручених один біля одного ланцюгів нуклеоти-дів. Нуклеотидів у молекулі ДНК — тисячі, їх молекулярна маса сягає десятків, мільйонів і більше. Послідовність нуклеотидів у полінуклеотидному ланцюзі ДНК теж різна. Це зумовлює високу видову специфічність білка. Нуклеотид складається із залиш-ка фосфорної кислоти і цукру (дезоксирибози в ДНК і рибози в РНК). До складу нуклеотиду входить також по одній з чотирьох азотистих основ — пуринових (аденін, гуанін) або піримідинових (цитозин, тимін). Нуклеотиди (мононуклеотиди) являють собою мономери, з яких побудовані нуклеїнові кислоти (полінуклеоти-ди). В останніх мононуклеотиди сполучені між собою через залишок фосфорної кислоти, яка відіграє роль зв'язувальної ланки.
Комплементарне сполучення азотних основ подвійної спіралі ДНК таке: аденін — тимін, гуанін — цитозин. Разом з тим ці відношення АТ/ГЦ у ДНК різних видів значно варіюють. Так, у вищих рослин і тварин вміст А—Т перевищує Г—Ц. У вірусів, бактерій і нижчих організмів описані великі коливання величини відношень А—Т/Г—Ц. У людини відношення А—Т/Г—Ц дорівнює 1,4/1. На підставі иритохімічних та інших даних у деяких ділянках хромосоми вміст А—Т або Г—Ц може бути значно підвищеним.
Просторову модель ДНК на підставі рентгеноструктурного аналізу запропонували в 1953 р. Джеймс Уотсон і Френсіс Крик. Ця модель добре пояснює фізико-хімічні і біологічні властивості ДНК, особливо механізм її репродукції в клітині. Основні принципи побудови моделі:
кожна молекула ДНК включає два довгі антипаралельні по-лінуклеотидні ланцюги, утворюючи подвійну спіраль, закручену спірально навколо центральної осі, а кожний нуклеотид розташований у площині, перпендикулярній до осі спіралі;
два ланцюги з'єднані водневими зв'язками, які утворюються між основними частинами молекул, котрі належать різним ланцюгам;
3) спіралізація та спарювання основ високо специфічне, оскільки відстань між вуглеводними компонентами двох спаре- них нуклеотидів постійна (1,1 нм); пуринові основи з'єднуються тільки з піримідиновими;
4) послідовність основ в одному ланцюгу визначає суворо комп- лементарну (відповідну) їй послідовність в іншій. Цими компле- ментарними парами, як зазначалось, є А—Т і Г—Ц (жал. б).
Властивість ДНК подвоюватися реалізується так: два ланцюги розділяються, і кожний з них слугує матрицею для синтезу

двох комплементарних (відповідних) ланцюгів. У результаті цього утворюються дві молекули ДНК, які мають цілком ідентичну молекулярну структуру. Генетична інформація закодована в послідовності основ ДНК. У зв'язку з тим, що молекула ДНК це довгий полімер, в якому можлива величезна кількість варіантів у розміщенні нуклеотидів, то за допомогою наявних чотирьох основ можна кодувати різну спадкову
інформацію.
Мал. 6. Фрагмент молекули ДНК Конструкція ДНК краще при- (схема)
Конструкція ДНК пристосована для здійснення її основної функції: зберігання постійного самовідновлення, самовідтворення (реплікацїї) та передачі на генетичному рівні інформації в клітині. Первинна структура РНК нагадує структуру ДНК, за винятком того, що РНК містить рибозу та урацил замість дезоксирибози та тиміну. Молекули РНК бувають односпіральними або, як і ДНК, двоспіральними. Є три типи РНК, які відрізняються за масою та іншими властивостями: рибосом на (рРНК), інформаційна, або матрична (ІРНК), розчинена, або транспортна (тРНК). Усі вони синтезуються в ядрі клітини і беруть участь у синтезі білків та ферментів.
Фізико-хімічні властивості цитоплазми. Фізико-хімічний стан цитоплазми залежить від різних станів навколишнього середовища. Під час дії різних чинників (хімічних, механічних, термічних та ін.) цитоплазма клітин переходить у стан паране-крозу (від грец. para — біля, песros — смерть).
Різні процеси метаболізму клітини здійснюються за участю ферментів, або ензимів. Ферменти (від лат. еп — усередині, zут-те —дріжджі, закваска) це біологічні каталізатори, тобто специфічні білкові речовини, які зумовлюють біохімічні перетворювання в процесі обміну речовин. За хімічною будовою ензими поділяють на прості (однокомпонентні) та складні (двокомпонентні). Прості ензими складаються з білка, а складні — з білка (апофермента) та небілкової частини (простатичної групи, або коферменте; від лат. со — разом, fermentum — закваска), яка включає вітаміни або атоми металів як реактивний центр ензимів. Дія ферментів специфічна: вони каталізують певні реакції.
Молекула будь-якого ферменту має один або декілька активних центрів, до яких прикріплюється молекула субстрату. Вони підходять один до одного подібно до того, як ключ підходить до замка. Тому кожний фермент діє лише на відповідний субстрат.
Багато ферментів добре вивчені, виділені, одержані в кристалічному вигляді та широко використовуються в біології, клінічній медицині, харчовій промисловості тощо. Характерним є те, що біологічні мембрани клітини являють собою своєрідні матриці, на яких розташовані ферментативні системи. Ці системи сприяють перетворенню хімічної енергії клітини на електричну і навпаки.
У клітину як структурну і функціональну одиницю живого проникають речовини. Закономірності надходження речовин у клітину зумовлюють явища проникливості клітинних мембран. Проникливість — здатність клітини поглинати і затримувати речовини, розчинені в середовищі, яка оточує клітинну мембрану, а також виділяти в це середовище речовини, вироблені в процесі життєдіяльності клітини. Проникливість здійснюється за рахунок наявності активного транспорту речовин, дифузії крізь біологічні мембрани, здатності білків цитоплазми адсорбувати речовини тощо. Проникливість клітинних мембран має важливе значення для обміну речовин, транспорту йонів, органічних речовин, води. З нею пов'язане також надходження в клітину лікарських препаратів, наркотичних засобів. Характерно, що проникливість стосовно одних речовин виражена досить пасивно, а стосовно інших — активно. Що вища розчинність, то швидше проникають речовини. Пасивне перенесення здійснюється проникненням крізь клітинну мембрану. Так найкраще проникають органічні молекули, що добре розчиняються в жирах.
Для клітин характерним є також явище дифузії (від лат. diffusio— поширення) — взаємопроникнення речовин та розподіл частинок унаслідок теплового руху. Дифузія здійснюється в результаті різниці концентрацій, у напрямку зменшеної концентрації речовин, і триває до моменту рівномірного їхнього розподілу. Дифузія відіграє важливу роль у процесі життєдіяльності клітини. Так, різна швидкість дифузії йонів крізь клітинні мембрани є одним із фізіологічних факторів, які впливають на вибірне накопичення елементів.
Поряд із дифузією велике значення для клітин має осмос (від грец. osmos — тиск) — одностороннє проникнення води крізь напівпроникну оболонку. Тиск, який виник на цій оболонці (мембрані), називається осмотичним. У хребетних і людини осмотичний тиск — постійний унаслідок регуляції.
Залежно від осмотичного тиску розчини поділяють на ізотонічні (у них тиск аналогічний тиску в клітині; ізотонічні розчини солей називають фізіологічними) та гіпертонічні (де осмотичний тиск вищий, ніж у клітинах).
Ріст і розвиток будь-якої клітини зумовлюються утворенням нової живої субстанції — білка. У процесі життєдіяльності клітини молекули білка зношуються і в ній синтезуються нові білки. Здатність до біосинтезу білків — одна з найважливіших особливостей клітини.
Біосинтез білка в клітині відбувається за принципом матричного синтезу за участю нуклеїнових кислот. Матричний шлях синтезу білка перебуває у взаємозв'язку з нуклеїновими кислотами, а саме — із ДНК, котра є матрицею (штампом), формою, де закодована первинна структура білка, яка синтезується під впливом цієї ДНК. Спочатку на структурі ДНК синтезуються молекули інформаційної, або матричної, РНК (ІРНК), які надалі переходять з ядра до цитоплазми, де вони з'єднуються з рибосомами і беруть участь у синтезі відповідних білків.
Генетична інформація, укладена в молекулі ДНК, спрямовує синтез того чи іншого білка. Так, білок крові — гемоглобін, синтезується клітинами печінки, інсулін — клітинами підшлункової залози тощо. Різні ділянки ДНК несуть відповідальність за синтез певних білків. Кожному білку відповідає визначений амінокислотний склад і їх точне послідовне розміщення.
Утворення білків з амінокислот починається з ферментативної активізації останніх. Під впливом комплексу ферментів амінокислоти перетворюються на їх активну стадію внаслідок взаємодії АТФ з нуклеїновими кислотами. АТФ, а в деяких випадках — і подібні їй макроергічні кислоти ЦТФ (цитидинтрифосфорна) і ГТФ (гуанінтрифосфорна) є тими акумуляторами, які безпосередньо забезпечують енергію як для активізації амінокислот, так і для інших етапів біосинтезу білка.
Біосинтез білка в клітині — складний хімічний багатоступінчастий процес, який відбувається у відповідному порядку на окремих структурних елементах клітини за участю різних систем ферментів. Особливість білкового синтезу клітин полягає в його винятково високій точності (безпомилковості). Генетично програмована структура білкових молекул зберігається з покоління в покоління. При цьому молекули синтезуються безліч разів у межах одного організму без виражених (у нормі) відхилень від заданої послідовності амінокислот.
У біосинтезі білка виділяють такі стадії:
активізація амінокислот, тобто перетворення їх на активні або реактивні форми;
з'єднання амінокислот з тРНК з утворенням складних комплексів між активними центрами амінокислот і тРНК;
транспортування комплексів амінокислот з тРНК до рибосом;
утворення поліпептидних ланцюгів на рибосомах;
від'єднання поліпептидного ланцюга від рибосом і утворення просторової структури білкової молекули.
Суть першої стадії полягає в тому, що під впливом відповідних ферментів (кожній амінокислоті відповідає специфічний фермент) амінокислоти вступають у взаємодію з донором енергії — АТФ. Реакція забезпечується наявністю деяких йонів — магнію, марганцю, кобальту. Утворюється продукт конденсації залишку амінокислоти із залишком аденілової кислоти (АМФ) — аміно-ациладенілат. Виділяється молекула пірофосфату. Амінокислота з'єднується із залишком аденілової кислоти за допомогою карбоксилфосфатного зв'язку. Цей зв'язок макроергічний, внаслідок якого енергія амінокислоти стає активною і переходить у подальші реакції утворення поліпептидного ланцюга в процесах біосинтезу білка. На початковій стадії процесу фермент утворює комплексне сполучення з АТФ, У результаті в молекулі АТФ відбувається послаблення зв'язку між залишками фосфатної кислоти. Потім комплекс взаємодіє з амінокислотою, унаслідок чого утворюється комплекс ферменту з аміноациладенілатом і вільний пірофосфат. Ферменти, які активізують амінокислоти, дуже поширені в природі. Для них характерна висока специфічність. Кількість ферментів дорівнює кількості амінокислот, які входять до складу білків.
Друга стадії полягає в перенесенні активних амінокислот і сполученні їх з ферментами та тРНК. Утворений при цьому комплекс амінокислот з відповідною специфічною для них тРНК має макроергічний зв'язок між залишком амінокислоти та тРНК.
Саме за рахунок енергії цього зв'язку і здійснюються подальші реакції. Сполучення активних амінокислот з тРНК відбувається за допомогою тих самих ферментних систем, які беруть участь і в активізації амінокислот. У вигляді комплексу з ферментом амі-ноациладенілат реагує з тРНК. Ця група ферментів, відповідно до їхньої функції, називається аміноацил-тРНК-синтетаза. Активізована амінокислота приєднується до кінцевої ділянки тРНК. Кінцева частина є структурою ЦЦА.
Третьою стадією біосинтезу білка є перенесення активованих амінокислот у комплексах з відповідними тРНК на нуклеопроте-їди рибосом клітини. Цей процес здійснюється також під впливом специфічних ферментів, кількість яких, мабуть, перевищує 20—60. Деякі амінокислоти транспортуються декількома тРНК. Так, для перенесення амінокислот валіну та лейцину існує по три різних тРНК, для процесу переносу амінокислот на рибосоми потрібна гуанозинтрифосфорна кислота (ГТФ) і система, яка синтезується АТФ.
На четвертій стадії на рибосомах синтезується специфічний білок. Вважають, що комплекс тРНК з активованими амінокислотами на поверхні рибосоми з'єднується водневими зв'язками з певною ділянкою інформаційної (матричної) РНК. Це відбувається за рахунок пуринових та піримідинових основ іРНК. На рибосомах клітини унаслідок дії відповідних ферментів, за участю нуклеопротеїдів цих клітинних структур, а також ІРНК та тРНК синтез білків здійснюється з використанням енергії.
Поліпептидний ланцюг відривається від рибосоми, утворюється на п'ятій стадії біосинтезу, після закінчення її синтезу. Заново утворений білок, відділений від рибосоми, переходить у цитоплазму і відповідно первинній структурі здобуває певну просторову конфігурацію, або макроструктуру.
Молекулярні основи механізму біосинтезу білка. Вони ґрунтуються на тому, що ДНК, сконцентрована переважно в хромосомах, безпосередньої участі в синтезі білка не бере.
Кодована в ДНК інформація про первинну структуру білка передається до рибосом, де здійснюється їх синтез. Незважаючи на те що рибосоми є апаратом синтезу білка, вони здатні забезпечувати цей процес (мал. 7). Утворення поліпептидного ланцюга з амінокислот потребує участі матричних (ІРНК) і транспортних (тРНК), а також АТФ і спеціалізованих ферментів та інших речовин. Отже, рибосоми можна назвати макромолекулярною ма-
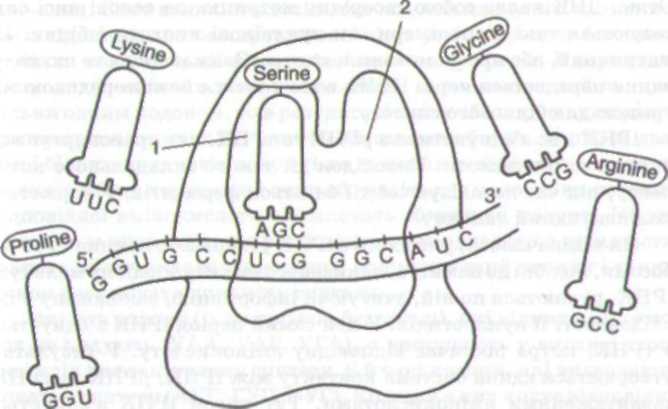
Мал. 7. Рибосома, на якій відбувається процес трансляції: 1 — пепти-дильний центр — Р-ділянка; 2 — аміноацильний центр — А-ділянка
шиною, що складається з багатьох підігнаних деталей і здатна до відбору компонентів, бере участь у трансляції та керує нею.
На поверхні ДНК унаслідок законів комплементарності може синтезуватися декілька молекул іРНК. Ділянки ДНК у функціональному відношенні неоднакові. Кожна з них складається з різної кількості триплетів і програмує синтез відповідного білка. Ці ділянки називають структурними генами. Деякі автори для поняття гена як функціональної одиниці пропонують термін "цитрон". Цитрон складається з триплетів, які називають кодонами. Відповідно кодонам ДНК на основі комплементарності формуються кодони іРНК. Тому й послідовність нуклеотидів цитрона ДНК відображається в послідовності нуклеотидів іРНК. Різниця лише в азотистих основах, де в іРНК замість тиміну, як у ДНК, міститься урацил. Саме комплементарність структур дає матриці змогу визначити нуклеотид і встановити його в належне місце.
Інформація про специфічність будови білкової молекули закладена в послідовності нуклеотидів молекули ДНК, яку можна назвати кодом. У зв'язку з тим, що спадкоємність пов'язана з біосинтезом специфічних білків, цей код іменується генетичним.
Отже, ДНК являє собою своєрідну матрицю, на основі якої синтезуються такі біополімери, як нуклеїнові кислоти і білки. Це матричний, або програмований, синтез. Зважаючи на те що кодування передається через іРНК, воно також є безпосередньою матрицею для білкового синтезу.
іРНК має з'єднуватися з рРНК та тРНК, яка транспортує відповідні амінокислоти. Унаслідок дії такого складального комплексу, під час каталізуючої дії багатьох ферментів, утворюється поліпептидний ланцюг.
Як відзначалося, рибосоми з іРНК утворюють ланцюг. Тут рибосоми, подібні до намиста, нанизаного на ниткоподібну молекулу іРНК, рухаються по ній, зчитуючи інформацію, закодовану в послідовності її нуклеотидів. У цей самий період іРНК з'єднується з тРНК, котра постачає відповідну амінокислоту. У результаті утворюється єдина система контакту між ІРНК, рРНК та тРНК, завантаженими амінокислотами. Тут кодон ІРНК з'єднується водневим зв'язком з кодоном (антикодоном) тРНК за принципом комплементарності (наприклад, А—Г, Ц—У). Отже, шляхи передачі інформації включають декілька етапів:
1) транскрипція — передача (переписування) інформації про послідовність нуклеотидів ДНК у послідовність нуклеотидів ІРНК;
2) транспортування іРНК від місця утворення до рибосом;
3) трансляція — перенесення інформації, закодованої в нукле- отидій структурі іРНК, на первинну структуру поліпептидного ланцюга білка.
Шляхи передачі інформації ґрунтуються на нуклеотидних кодонах. До складу молекули ДНК входять десятки тисяч комбінацій чотирьох різних нуклеотидів, які відрізняються один від одного своїми азотистими основами. Положення в молекулі ДНК однієї окремо взятої азотистої основи не може визначати положення окремої амінокислоти в молекулі білка. Азотистих основ у молекулі ДНК усього чотири, а амінокислот у молекулі білка — 20. Тому лише комбінації з трьох основ відповідають необхідним вимогам. Кількість можливих комбінацій по три з чотирьох дорівнює 43, тобто 64. Положення в молекулі білка кожної з 20 амінокислот визначається групою з трьох азотистих основ молекули ДНК.
Таким чином, виникає можливість кодування однієї амінокислоти декількома кодонами. Це явище має визначне біологічне значення. Саме наявність декількох кодонів для однієї амінокислоти підвищує динамічну гармонійність усього процесу біосинтезу білка внаслідок дії різних факторів на організм. Наприклад, якщо певна амінокислота включається в поліпептидний ланцюг тільки одним кодоном, то в результаті зміни або порушення цього триплету змінюється і біосинтез визначених білків. Якщо ж два, три і більше триплетів кодують включення певної амінокислоти, то якщо й буде порушений один або навіть два кодони, включення відповідної амінокислоти забезпечать збережені кодони. Не виключена можливість, що різні кодони з неоднаковою швидкістю можуть включати амінокислоти в поліпептидний ланцюг і таким чином регулювати динаміку синтезу.
Існують кодони (їх називають безглузді), які ніяких амінокислот не кодують (УАА, УАГ, УГА), а виступають у вигляді стоп-сигналів для закінчення синтезу. Є й такі кодони, які визначають початок синтезу (АУГ, ГУГ, УУГ). Кожний з них кодує відповідну амінокислоту і не відрізняється від інших кодонів. їхня роль залежить від положення їх у молекулі іРНК. Якщо вони розташовані на початку або в кінці молекули, то з ними тРНК не з'єднується, а відбувається зв'язок з тією тРНК, яка несе амінокислоту, з якої розпочинається синтез.
Уже розкрито не тільки склад кодонів, а й порядок чергування мононуклеотидів у них. Послідовне розміщення таких кодонів у іРНК або в їхніх комплексах з рибосомними нуклеотидами відіграє важливу роль в утворенні в процесі біосинтезу певного білкового ланцюга.
Отже, початком біосинтезу білка є гетеросинтетична функція гена, транскрипція — синтез іРНК — і перехід ІРНК у цитоплазму до рибосом (мал. 8).
У наслідок
активізації
амінокислот встановлю-
ється
їх зв'язок з тРНК
і утворюється
склад-
ний біологічний комп-
лекс:
амінокислота —
АТФ — специфічний
фермент
— тРНК. Потім
відбувається перенесен-
ня
амінокислот за допомогою тРНК на
рибосому.-
Мал.
8.
Фрагмент
молекули
наслідок
активізації
амінокислот встановлю-
ється
їх зв'язок з тРНК
і утворюється
склад-
ний біологічний комп-
лекс:
амінокислота —
АТФ — специфічний
фермент
— тРНК. Потім
відбувається перенесен-
ня
амінокислот за допомогою тРНК на
рибосому.-
Мал.
8.
Фрагмент
молекули
ДНК під час транскрипції
Амінокислоти за допомогою відповідних ферментів з'єднуються між собою. Виникає первинна структура білка з наступним завершенням формування вторинної, третинної та четвертинної структур. Таким чином, проблема біосинтезу білків як процесу самооновлення на молекулярному рівні може бути представлена так: ДНК (ген) — ІРНК — поліпептид — специфічний білок — ознака.
Спадкова інформація може передаватися не тільки від ДНК до РНК, а й у зворотному порядку за допомогою спеціального ферменту зворотної транскриптази. Ця особливість клітини розкрита на моделі онкогенних вірусів і може відігравати важливу роль у процесах онтогенезу та явищах імунітету.
Біологічний процес синтезу білка досить складний і здійснюється внаслідок реалізації ауто- та гетеросинтетичної функції гена. У результаті аутосинтетичної функції інформація, властива даному виду, може передаватися з покоління в покоління. На підставі ауто- та гетеросинтетичної функції і відбувається самооновлення клітини, забезпечується реалізація записаної в ній інформації. Самооновлення здійснюється завдяки обміну речовин клітини із зовнішнім середовищем у процесі саморегуляції та самовідтворення. Клітина, здійснюючи саморегуляцію, підтримує певну стабільність внутрішньо-клітинного середовища. В основі цього явища лежать фізико-хімічні процеси, що відбуваються на молекулярному рівні саморегуляції.
У вищих організмів генетичний контроль і саморегуляція складніші, ніж у мікроорганізмів. Незважаючи на те що геном у різних клітинах і тканинах одного організму однаковий, у них синтезуються різні білки (наприклад, пепсин — у слизовій оболонці шлунка, гемоглобін — в еритроцитах та ретикулоцитах тощо). Це можна пояснити блокуванням більшої частини генів у результаті диференціювання, яке відбувається за участю гістонів та інших білків хроматину.
Проте з гістонами пов'язана особлива, раніше невідома різновидність РНК. На підставі цього запропоновано гіпотезу, за якою саме ІРНК, а не гістонам належить специфічна дія. Саме така РНК блокує оператор, вступаючи з ним у комплементарні взаємовідносини. Існують й інші механізми саморегуляції діяльності, що здійснюються в клітині. Усе ж таки провідна роль у саморегуляції діяльності клітини належить, як зазначалось, її генетичному апарату.
І
Життєвий цикл клітини. Поділ клітини
Поділ клітини відбувається після подвоєння власного генетичного матеріалу, маси клітини та її компонентів. Ці передумови поділу в свою чергу відбуваються в інтерфазі клітинного циклу. Клітинний (життєвий) цикл включає народження, поділ і смерть клітини.
Нова клітина, згідно із клітинною теорією, утворюється лише за допомогою поділу. У амеби ротової та інших одноклітинних організмів клітинний цикл триває впродовж усього життя, тобто у них клітинний цикл збігається з життєвим. Наприклад, тривалість клітинного циклу амеби-протей — 25—35 год — тобто життя становить також цей час.
Клітини багатоклітинних організмів протягом життя розмножуються певну генетично зумовлену кількість разів. Клітинний цикл таких організмів повторюється відповідну кількість разів. Кожний цикл складається з чотирьох періодів: пресинтетично-го, або постмітотичного (О,), синтетичного (8), постсинтетично-го, або премітотичного (С2)» і мітозу. Перший, пресинтетичний, період клітинного циклу проходять усі клітини. Різні клітини перебувають у цьому періоді різний час, водночас для нервових еоіітин це єдиний період. Клітини ростуть, накопичують різні білки. Гени виконують гетеросинтетичну функцію. У клітині нагромаджуються багаті на енергію речовини, нуклеотиди, амінокислоти, ферменти. На цьому етапі клітина готується до подвоєння генетичного матеріалу. До наступного, синтетичного, періоду переходять лише ті клітини, що минули точку рестрикції, яка супроводжується наявністю в клітині нестабільного білка у певній концентрації. Відбувається реплікація ДНК і подвоєння матеріалу хромосом; синтез білка та РНК.
У синтетичному періоді підвищується активність ферменту ДНК-полімерази, а внаслідок аутосинтетичної функції гена відбувається подвоєння генетичного матеріалу — реплікація молекули ДНК. Синтез ДНК починається одночасно в кількох місцях хромосоми. Можливо, ці "стартові" точки реплікації з'єднані внутрішньою поверхнею ядерної мембрани. Ділянка хромосоми, розміщена між двома сусідніми "стартовими" точками, є одиницею реплікації ДНК; вона називається репліконом. У кінці синтетичного періоду кожна хромосома стає двохроматидною. Наприклад, у клітинах людини буде 92 молекули ДНК та 46 двохроматидних хромосом.
Третій, постсинтетичний, період супроводжується синтезом білків веретена поділу та підготовкою клітини до мітозу. Ядра збільшуються. Центросома перетяжкою ділиться навпіл.
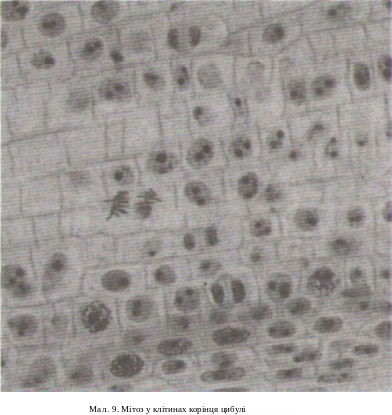 Четвертий
період — мітоз. Непрямий поділ, або
мітоз, характеризується процесами,
які відбуваються в ядрі і клітині в
цілому
(мал.
9).
Генетичний
матеріал клітини, що подвоївся в
синтетичному періоді інтерфази,
завдяки мітозу рівномірно розподіляється
між двома новими дочірніми клітинами.
Материнська і дочірня клітини мають
однакову кількість хромосом та ідентичні
Четвертий
період — мітоз. Непрямий поділ, або
мітоз, характеризується процесами,
які відбуваються в ядрі і клітині в
цілому
(мал.
9).
Генетичний
матеріал клітини, що подвоївся в
синтетичному періоді інтерфази,
завдяки мітозу рівномірно розподіляється
між двома новими дочірніми клітинами.
Материнська і дочірня клітини мають
однакову кількість хромосом та ідентичні

генетичні програми. Якщо мітоз відбувся без порушень, то соматична клітина людини має 46 однохроматидних хромосом і 46 молекул ДНК. Для мітозу характерні послідовні морфологічні та фізіологічні зміни, які втягують у процес ядро, цитоплазму та її органели.
Перша фаза мітозу — профаза (мал. 10, Б). Клітина округлюється, цитоплазма стає більш в'язкою, тургор її підвищується, збільшується поверхневий натяг клітини. Центросоми, які розділилися, розходяться все далі по довгій осі клітини до її полюсів. Між центросомами відбувається формування ахроматинового веретена, яке складається з пружних білкових ниток. Ці нитки за своїм розміщенням нагадують силові лінії магнітного поля. У період профази хромосоми помітно ущільнюються, потовщуються, укорочуються. У такому стані вони добре проглядаються в світловому мікроскопі. Відбувається розчинення ядерців. Розпізнавальною ознакою закінчення профази є розчинення оболонки ядра, у результаті чого хромосоми опиняються в загальній масі цитоплазми та каріоплазми.
Друга фаза — метафаза (мал. 10, В). Хромосоми розміщуються в одній площині по екватору. Таке характерне розміщення їх називають екваторіальною, або метафазною, платівкою. На цій стадії визначається кількість і форма хромосом, особливо при розгляданні метафазної платівки з полюсів поділу клітини.
Метафазну платівку використовують для визначення каріотипу. Сформоване ахроматинове веретено, на відміну від хромосом, не забарвлюється барвниками. Нитки ахроматину являють собою пучки мікротрубочок, які складаються головним чином з білка, який нагадує скорочувальний білок м'язових волокон. Розташовані ахроматинові нитки так, що в сукупності утворюють фігуру, подібну до веретена. Сумісно з центріолями ахроматинове веретено утворює мітотичний апарат клітини. Для цитоплазми в цей період характерна найменша в'язкість. Хромосоми своїми нерозді-леними центромерами розміщуються в екваторіальній площині, перпендикулярно до осі веретена. Центромера кожної хромосоми перебуває точно в екваторіальній площині, решта ж тіла хромосоми може бути і поза нею.
Третя фаза — анафаза {мал. 10, Г). Хромосоми розчіплюються, і сестринські хромосоми (хроматиди) розходяться до різних полюсів. На цій стадії скорочуються тягнучі нитки веретена за рахунок енергії АТФ та відштовхуються дочірні хромосоми. Відштовхуються одна від одної насамперед центромерні ділянки, а потім розходяться до полюсів центромерами вперед і самі сестринські хромосоми. Розходження хроматид в анафазі починається одночасно і закінчується дуже швидко.
Четверта фаза — телофаза (мал. 10, Д). Відбувається деспіра-лізація дочірніх хромосом, і вони втрачають видиму індивідуальність. Утворюється оболонка ядра, а потім і ядро. Реконструкція ядра здійснюється в зворотному порядку. Відновлюються ядерця в тій кількості, яка була у вихідній формі. Формується мембрана, відокремлюючи дочірні клітини одну від одної. Відбувається розподіл органел між новими клітинами. У період телофази повністю відокремлюються дочірні клітини шляхом поділу цитоплазми — цитокінезу.
Є випадки множинної реплікації молекул ДНК без наступного поділу клітин (цитотомії). Таке явище називається ендомітозом. Ендомітоз є джерелом утворення поліплоїдних клітин, в яких набір хромосом збільшений у кратну кількість разів. Таке явище характерне, наприклад, для печінки, де виявляються клітини, які
мають десятки і сотні ядер. Нарівні з ендомітозом спостерігається і явище політенії (від грец.роіу — багато, іаепіа — нитка). Це випадок ендомітозної поліплоїдії, коли кількість хромонем у хромосомах збільшується, але хромосоми не розчіплюються, а значно стовщуються, досягаючи величезних розмірів (політенні хромосоми). У випадках політенії випадають усі фази мітотичного циклу, крім основної — репродукції первинних ниток-хромосом.
Загальна тривалість життєвого циклу, включаючи і період мітозу, різна — від 24 до 72 год і більше, залежно від типу тканин, умов життя клітини і віку організму. Найкоротший мітотичний цикл (11 год) спостерігається в епітеліальних клітинах дванадцятипалої кишки. В епітелії рогівки ока тривалість циклу становить 72 год.
З усього клітинного циклу найтривалішим є пресинтетичний період, на який припадає близько половини всього часу. Пост-синтетичний період і мітрз порівняно короткочасні, тривалість їх 1—4 год, а синтетичного — 8—9 год. При цьому зазначений термін характерний для клітин як з коротким (24 год), так і з тривалим (72—114 год) мітотичним циклом. Фізичний ріст людини зумовлений нормальним процесом ділення клітин. Майже 50 послідовних поділів утворюють величезну кількість клітин, які складають тіло дорослої людини. Клітинний цикл — саморегулівний процес. Кожний подальший його етап здійснюється за рахунок попередньої стадії і, у свою чергу, зумовлює початок нової стадії. Мітотична активність залежить не тільки від самих клітин, а й від умов життя, які впливають на хід циклу.
В організмі спостерігається добова ритмічність мітозу. Якщо орган функціонує напружено, то мітотична активність його клітин значно знижується. На мітотичну активність впливають гормони: наприклад, гормон надниркової залози гальмує мітоз; гормони яєчника підвищують мітоз клітин грудної залози та матки, а гормон гіпофіза — мітоз клітин щитоподібної залози. Установлено, що у відмерлих клітин виділяються особливі речовини (ракові гормони), які стимулюють клітинний цикл живих клітин. Що більше клітин відмирає, то інтенсивніше відбувається поділ залишених. Мітотична активність клітин може змінюватися під впливом нервової системи. Після денервації якого-небудь органа порушується добова ритмічність мітозу. У період мітозу синтетичні процеси в клітині різко скорочуються, і протягом більшої частини клітинного циклу клітина зайнята підготовкою до самовідтворення.
Порушення мітозу призводить до появи поліплоїдів. Цей термін запропонував у 1916 р. Г. Венклер, який розумів під поліплоїдією змінену кількість хромосом. Згідно з цитологічними дослідженнями, більшість видів покритонасінних рослин — це поліплоїди. Гаплоїдне число хромосом виду генетики називають основним числом. У процесі еволюції і експериментальних досліджень поліплоїди виникають переважно в результаті порушення мітозу, а інколи мейозу.
Анеуплоїди — це поліплоїди, диплоїдні клітини яких мають в основному наборі збільшену або зменшену кількість хромосом на одну або декілька. Моносоміки — це анеуплоїди, у яких з пари гомологічних хромосом недостатня одна з гомологічних хромосом будь-якої пари (2л-1). Трисоміки —- це анеуплоїди, у яких одна гомологічна пара має три хромосоми (2л+1). Більшість анеупло-їдних зигот гинуть на ранніх стадіях ембріогенезу. Прикладами життєздатних анеуплоїдій у людини є хромосомні хвороби: хвороба Дауна — у 94 % випадків це трисомія 21-ї пари; синдром Патау — трисомія 13-ї пари хромосом; синдром Едвардса — трисомія 18-ї пари хромосом; синдром Шерешевського—Тернера — моносомія 23-ї пари хромосом.
Амітоз — прямий поділ клітини. Особливістю амітозу є те, що ДНК перед поділом подвоюється, але хромосоми не стають помітними, ахроматинове веретено не утворюється. Відсутність веретена поділу призводить до нерівномірного розподілу хромосом. Ядро клітини ділиться навпіл перетяжкою. Амітоз відбувається при поділі патологічно змінених клітин, а також у спеціалізованих клітинах печінки.
Мейоз — поділ ядра клітини, який призводить до зменшення вдвічі кількості хромосом диплоїдної клітини. Мейоз властивий організмам, які розмножуються статево. Завдяки мейозу статеві клітини, або гамети, завжди гаплоїдні. Мейоз складається з двох послідовних поділів клітини. Під час першого — редукційного періоду кількість хромосом зменшується вдвічі; після другого поділу кількість хромосом залишається гаплоїдним, а хромосоми з двохроматидних стають однохроматидними. У результаті цього кількість молекул ДНК відповідає кількості однохроматидних хромосом, або сестринських хромосом.
Перший і другий мейотичні поділи складаються з чотирьох фаз: профази, метафази, анафази і телофази. Редуплікація ДНК відбувається тільки перед першим поділом.
Профаза І мейотичного редукційного поділу має п'ять стадій: лептонему, зигонему, пахінему, диплонему і діакінез. У лептоне-мі ядро збільшується, хромосоми двохроматидні і мають вигляд тонких деспіралізованих ниток. У зигонемі парні гомологічні хромосоми наближуються і всіма ділянками зливаються. Утворюються біваленти, кожний бівалент складається з чотирьох хрома-тид. Злиття двох гомологічних хромосом називають кон'югацією. У пахінемі біваленти вкорочуються і стовщуються. На цій стадії між ідентичними ділянками гомологічної пари хромосом відбувається перехрест і обмін — кросинговер. У диплонемі продовжується вкорочення і стовщення, яке призводить до відштовхування сестринських хроматид. У діакінезі хромосоми спіралізовані, оболонка ядра руйнується, починається наступна фаза.
Метафаза І: гомологічні хромосоми розташовані попарно в екваторіальній площині клітини. Кожна хромосома двохрома-тидна.
Анафаза І: з кожної пари гомологічних хромосом до протилежних полюсів клітини розходиться по одній хромосомі. Так відбувається редукція (зменшення вдвічі) кількості хромосом. Розподіл хромосом — явище випадкове і зумовлює появу різних гамет.
Телофаза І: навколо хромосом на полюсах клітини формується ядерна оболонка, потім відбувається цитокінез. Кожна з двох дочірніх клітин має гаплоїдний набір хромосом (п) та подвоєну кількість ДНК, тому що хромосоми мають дві хроматиди.
Після короткого періоду — інтеркінеза (проміжку між кінцем першого і початком другого мейотичного поділу) — починається другий мейотичний поділ.
Другий мейотичний поділ подібний до мітозу. Профаза II супроводжується: спіралізацією двохроматидних хромосом, руйнацією ядерної оболонки і зникненням ядерця, формуванням ахроматинового веретена поділу. Метафаза II характеризується наявністю сформованого веретена поділу; центромери хромосом розміщуються в один ряд по екватору. До хромосом приєднуються нитки ахроматинового веретена.
Анафаза II супроводжується поділом центромер кожної хромосоми і розходженням хроматид до полюсів клітини. Телофаза II характеризується деспіралізацією хромосом, розчиненням веретена поділу і формуванням ядерця та ядерної оболонки. Потім відбувається цитокінез.
Після II мейотичного поділу утворюється чотири гаплоїдні клітини; усі хромосоми однохроматидні. У людини мейоз закінчується утворенням гамет, що мають 23 хромосоми і 23 молекули ДНК(ІпІС).
Генетичне значення мейозу полягає у тому, що завдяки йому:
підтримується сталість диплоїдного набору хромосом у кожного виду організму. У людини соматична клітина має 46 хромосом;
незалежно розходяться хромосоми в анафазі І та хроматиди в анафазі II. Це один із механізмів забезпечення генетичної перекомбінації хромосом матері і батька. Випадкова пере-комбінація забезпечує генетичну різноманітність гамет;
обмін генами між хроматидами гомологічних хромосом, або кросинговер, забезпечує генетичну перекомбінацію. Тому кожна гамета організму є генетично унікальною і неповторною.
РОЗДІЛ 2
Молекулярні основи спадковості. Реалізація спадкової інформації
Характеристика нуклеїнових кислот: ДНК і РНК, просторова організація, видова специфічність, роль у зберіганні та передачі спадкової інформації
Уперше нуклеїнові кислоти виявив Ф. Мішер у 1869 р.
Переконливі докази того, що саме з ДНК пов'язана передача спадкової інформації, отримані при вивченні вірусів. Проникаючи в клітину, вони вносять у неї лише нуклеїнову кислоту з дуже невеликою кількістю білка, а вся білкова оболонка залишається поза клітиною. Отже, введена в клітину ДНК передає генетичну інформацію, необхідну для утворення такого самого біологічного виду. Виявлено, що чиста нуклеїнова кислота вірусу тютюнової мозаїки може заразити рослину і спричинити типову картину захворювання. Більше того, вдалося штучно створити вегетативні "гібриди" з вірусів, у яких білковий футляр належить одному виду, а нуклеїнова кислота — іншому. У таких випадках генетична інформація "гібридів" завжди з точністю відповідала тому вірусу, нуклеїнова кислота якого входила до складу "гібриду".
Вагомі докази ролі ДНК у передачі спадкової інформації отримані також в експериментах на мікроорганізмах завдяки явищам трансформації, трансдукції і кон'югації.
Трансформація (від лат. transformatiо — перетворення) — включення чужорідної ДНК у геном клітини-хазяїна, що призводить до зміни її структурних і функціональних властивостей. Спадкова інформація від однієї клітини до іншої передається за допомогою ДНК клітини-донора. Явище трансформації було виявлено в дослідах англійського мікробіолога Гріффітса (1928).
Трансдукція (від лат. transductio — переміщення) полягає в тому, що віруси, залишивши бактеріальні клітини, в яких вони паразитували, можуть захоплювати частину її ДНК і, потрапив
ши в нові клітини, передають новим хазяїнам властивості попередніх. Це явище вперше було відкрито в дослідженнях щодо зараження бактерій вірусами.
Кон'югація (від лат. сопіjugatіо — з'єднання) — це передача генетичного матеріалу від однієї бактерії до іншої шляхом утворення цитоплазматичного містка, переміщення частини ДНК та її інтеграція з геномом клітини-реципієнта.
Будова молекули ДНК. Згідно з моделлю Уотсона і Крика, макромолекула ДНК — це два довгі полімерні ланцюги, що складаються з мономерів дезоксирибонуклеотидів, тісно з'єднаних між собою. Нитки ДНК з'єднуються водневими зв'язками між азотистими основами двох ланцюгів і утворюють подвійну спіраль ДНК. Пуринові та піримідинові основи взаємодіють одна з одною. Аденін одного ланцюга двома водневими зв'язками з'єднується з тиміном іншого ланцюга, а гуанін — трьома водневими зв'язками з цитозином. Таке сполучення азотистих основ забезпечує міцний зв'язок обох ланцюгів. Два полінуклеотидні ланцюги ДНК антипаралельні. Тобто, 5'-кінець одного ланцюга з'єднаний із 3'-кінцем іншого, і навпаки. Генетична інформація записана послідовністю нуклеотидів у напрямку від 5'-кінця до З'-кіиця. Така нитка називається "змістовною", саме тут розташовані гени (матричний ланцюг). Другий ланцюг у напрямку 3'—5' вважається "антизмістовним". Він необхідний як "еталон" збереження генетичної інформації і набуває значення в процесах реплікації та репарації.
Два довгі антипаралельні полімерні ланцюги, що складаються із дезоксирибонуклеотидів, міцно з'єднані між собою водневими зв'язками. У результаті цього утворюється подвійна спіраль, закручена навколо центральної осі.
Діаметр подвійної спіралі ДНК становить 2 нм, відстань між двома завершеними витками — 3,4 нм. У кожний виток входить 10 пар нуклеотидів. Відстань між сусідніми основами — 0,34 нм.
ДНК — це полімер, мономерами якого є нуклеотиди. Нуклео-тид складається з: 1) азотистої основи; 2) моносахаридів дезоксирибози; 3) залишку фосфорної кислоти.
Азотисті основи бувають двох типів: пуринові — аденін (А) і гуанін (Г) та піримідинові — тимін (Т) і цитозин (Ц).
До складу молекули ДНК входять чотири типи нуклеотидів: аденінові, гуанінові, тимінові, цитозинові. З'єднання нуклеотидів одного ланцюга в молекулі ДНК відбувається в результаті взаємодії фосфату одного нуклеотиду з гідроксильною групою дезоксирибози іншого. У результаті утворюється фосфодіефір-иий зв'язок, що об'єднує нуклеотиди в довгий ланцюг. Скелет ланцюга складається з молекул фосфату і пентоз, які чергуються між собою. Полінуклеотидний ланцюг синтезується за участю ферменту ДНК-полімерази. Цей фермент приєднує фосфатну групу одного нуклеотиду до гідроксильної групи дезоксирибози наступного.
Комплементарність пар основ. Два полінуклеотиднї ланцюги ДНК не ідентичні, але вони комплементарні один одному. Це пов'язано зі строгою відповідністю основ одного ланцюга основам іншого. Відстань між двома ланцюгами ДНК становить 2 нм, тому вміщується тільки одна пара комплементарних нуклеотидів (А—Т або Г—Ц). Тільки аденін і тимін, а також гуанін і цитозин мають відповідні просторові структури для утворення водневих зв'язків. Концепція специфічзого зв'язування пар основ свідчить, що аденін в одному ланцюгу відповідає тиміну в іншому, а гуанін має навпроти себе цитозин в іншому ланцюгу. Таким чином, два ланцюги ДНК комплементарні один одному.
Колінеарність— властивість, що зумовлює відповідність між послідовностями триплетів нуклеотидів (кодонів) нуклеїнових кислот і амінокислот поліпептидних ланцюгів. Тобто послідовність амінокислот білка, в якій відповідні кодони розташовуються в гені. Це означає, що положення кожної амінокислоти в поліиептидному ланцюгу білка залежить від положення відповідного триплету в гені. Генетичний код вважається колінеарним, тобто кодони нуклеїнових кислот і відповідні їм амінокислоти білка розташовані в однаковому лінійному порядку.
Явище колінеарності доведено експериментально. Так, установлено, що серпоподібноклітинна анемія, за якої порушена будова молекули гемоглобіну, зумовлена зміною одного нуклеотиду в його гені, що призводить до заміни однієї амінокислоти на іншу. Гіпотезу про те, що послідовність нуклеотидів у гені визначає послідовність амінокислот білка, висунув Г.А. Гамов (1954). Дані про колінеарність поліпептидів підтвердили її. Завдяки концепції колінеарності можна визначити порядок нуклеотидів усередині гена і в ІРНК, якщо відомий амінокислотний склад поліпептидів, і навпаки, визначивши склад нуклеотидів ДНК, можна передбачити амінокислотний склад білка. Цей принцип використовується в методах молекулярної біології. Із цієї концепції видно, що зміна порядку нуклеотидів усередині гена (його мутація) призводить до зміни амінокислотного складу білка.
Видова специфічність ДНК. За співвідношенням А+Т і Г+Ц представники різних видів відрізняються між собою, причому у тварин переважає пара А+Т, а в мікроорганізмів співвідношення А+Т і Г+Ц однакове. Ці явища використовують як один із генетичних критеріїв визначення виду. У цьому полягає видова специфічність ДНК.
Просторова організація ДНК. Молекула ДНК може існувати в різній конфігурації залежно від навколишніх умов. Відомо декілька форм ДНК:
а) В-форма — має стандартну структуру відповідно до моделі молекули Уотсона і Крика, у нормальних фізіологічних умовах є основним структурним типом;
б) А-форма — виявлена у зневодненому середовищі, при висо- кому вмісті калію і натрію. Така ДНК має дещо змінену спіралі- зацію;
в) с-форма — має менше основ на один виток, а отже, інші — фізичні характеристики;
г) Z-форма — на відміну від інших форм закручена вліво. Дея- кі форми при зміні фізіологічних умов можуть переходити одна в одну, що додатково регулює роботу генів.
Знаючи структуру ДНК, можна зрозуміти суть багатьох молекулярно-генетичних процесів. Так, у молекулі ДНК виділяють структури: первинну — послідовність нуклеотидів у ланцюгу; вторинну — два комплементарні ланцюги, з'єднані водневими зв'язками; третинну — тривимірну спіраль.
Зазначимо, що: а) геометрія спіралі ДНК залежить від послідовності нуклеотидів; б) значна частина ДНК не кодує білків або РНК; в) кожний ген — це складова функціонально-активна одиниця, призначена для регульованого синтезу РНК.
Рибонуклеїнові кислоти. Уся спадкова інформація зберігається в молекулі ДНК. Передачу спадкової інформації від ДНК у цитоплазму здійснюють рибонуклеїнові кислоти як посередники. Взаємовідносини ДНК, РНК і білків можна представити у вигляді схеми: ДНК — РНК — білок.
У цьому випадку один з ланцюгів ДНК є матрицею для молекул РНК, які, зокрема, є матрицями синтезу білків або входять до складу рибосом чи переносять амінокислоти.
РНК складаються з одного ланцюга і мають вигляд довгих перозгалужених полімерних молекул. Одноланцюгові РНК можуть утворювати подвійні спіралі, якщо різні частини ланцюга мають антипаралельні комплементарні сегменти, пов'язані один і одним. У деяких вірусів РНК є носієм спадкової інформації за відсутності ДНК. Частина РНК мають каталітичну активність на певні клітинні процеси. РНК — це полімер рибонуклеоти-дів, які складаються з фосфорної кислоти, рибози та азотистих основ (аденіну, гуаніну, цитозину, урацилу). Рибоза разом із залишками фосфорної кислоти утворює скелет молекули, на якому розміщені азотисті основи. Усі різновиди РНК синтезуються на молекулах ДНК за участю ферментів РНК-полімераз за принципом комплементарності. При цьому в синтезованій молекулі РНК аденін ДНК комплементарний урацилу РНК, а гуанін — цитозину. Якщо вміст ДНК у клітинах постійний, то вміст РНК дуже коливається залежно від типуклітини, інтенсивності метаболізму і синтезу білків.
Що стосується молекул РНК, то вони мають багато спільного зі структурою ДНК, але відрізняються низкою ознак: а) вуглеводом РНК є рибоза; б) РНК не містить тиміну, його місце в молекулі займає урацил; в) РНК — одноланцюгова молекула; г) правила Чаргаффа не діють.
Типи РНК. Розрізняють три типи РНК залежно від розміру, структури і функції молекул, характерних як для прокаріотів, так і для еукаріотів.
Інформаційна РНК (іРНК). На певних ділянках ДНК утворюються її молекули (вони називаються структурними генами) у вигляді комплементарної копії ділянки одного з її ланцюгів. Вони несуть закодовану інформацію первинної структури білків у цитоплазму, де прикріплюються до рибосом і реалізують цю інформацію.
Під час синтезу поліпептидів (білків) роль матриці виконує інформаційна РНК, тому її називають також матричною. Матрична РНК є шаблоном, на якому будуються поліпептиди відповідно до закладеної генетичної інформації. Зазвичай, вона несе інформацію про синтез тільки однієї молекули білка — це моноцистронна іРНК. Іноді вона містить декілька цистронів, розташованих поряд, для різних білків і відома під назвою поліцистронна ІРНК.
Інформаційна РНК містить інформацію про порядок розміщення амінокислот у синтезованому білку. Розміщення амінокислот кодується чіткою послідовністю нуклеотидів у молекулі ІРНК (генетичний код). Кожній амінокислоті відповідає свій триплет нуклеотидів (кодон). Молекули ІРНК складаються з 300—3000 нуклеотидів. Вони становлять 0,5—3,0 % маси всіх РНК клітини. Інформаційна РНК утворюється в ядрі у вигляді незрілої про-іРНК, яка містить і неінформативні послідовності нуклеотидів (інтрони). У результаті процесингу (вирізання ін-тронних ділянок) вона "дозріває" і надходить у цитоплазму, де відразу приєднується до рибосом. Проте іноді ІРНК може накопичуватися в клітинах, зв'язуватися зі спеціальними білками, що "консервують" її, утворюючи інформосоми. У такому вигляді інформація може тривалий час зберігатися в клітинах. Поштовхом для її використання є фізіологічні зміни в клітині, що призводять до активації синтезу білка. Наприклад, в овоциті накопичується багато інформосом, а їхня ІРНК починає функціонувати тільки після запліднення.
Транспортна РНК (тРНК). Молекули тРНК утворюються на спеціальних генах. Транспортні РНК короткі, однониткові, мають форму листка конюшини завдяки комплементарному сполученню основ на різних ділянках ланцюга, складаються з невеликої кількості нуклеотидів (75—90). Від загальної маси РНК на тРНК припадає близько 10—15 %. Молекули тРНК переносять до місць синтезу білків тільки відповідні їм амінокислоти з цитоплазми. Кожній амінокислоті відповідає своя тРНК унаслідок особливостей нуклеотидної послідовності та просторової структури. Молекули тРНК мають чотири важливі ділянки: а) транспортну; б) антикодон; в) ділянку приєднання ферменту; г) ділянку зв'язування з рибосомою.
До транспортної ділянки приєднується специфічна амінокислота. Вона утворена двома комплементарними кінцевими ділянками РНК, З'-кінець якої складається із семи пар основ. Він довгий і формує одноланцюгову ділянку, шо закінчується послідовністю ЦЦА з вільною ОН-групою. До цієї групи приєднується амінокислота, що транспортується.
Антикодон складається з п'яти нуклеотидів. У центрі — три специфічні рибонуклеотиди (триплет). Азотисті основи анти-кодона мають комплементарний триплет на ланцюгу ІРНК. Цей триплет називається кодоном. У період синтезу білка антикодон знаходить відповідний йому кодон на ІРНК і тимчасово приєднується до нього водневими зв'язками.
Розділ 2
53
Ділянка приєднання ферменту — це спеціальна частина молекули тРНК для специфічного зв'язування з ферментом аміноацил-тРНК-синтетазою, що каталізує приєднання амінокислоти до рибосоми.
Рибосомальна РНК (рРНК). Рибосомальна РНК утворюється на спеціальних генах ДНК в ядерці. Це велика одноланцюгова розгалужена молекула, що включає 3000—5000 нуклеотидів. Із ІагальноІ маси РНК на її частку припадає до 90 % . У каріоплазмі і»ГІІК і різні білки об'єднуються у співвідношенні 1:1 для утворення малих і великих субодиниць рибосом.
Рибосомальна РНК утворює структурний каркас рибосоми, їй належить важлива роль у процесі синтезу білків. Рибосомальна РНК .шбезпечуе зв'язування іРНК з рибосомами за допомогою певних послідовностей нуклеотидів. Таким чином, установлюються початок і рамка зчитування інформації з ІРНК. Багато білків рибосом виконують не тільки структурну, а й ферментативну функцію.
Отже, чотири різновиди нуклеїнових кислот мають багато спільного в будові, але виконують різні функції.
Реплікація ДНК
Унікальна властивість молекули ДНК подвоюватися перед поділом клітини називається реплікацією. Ця властивість зумовлена особливістю будови молекули ДНК, що складається з двох комплементарних ланцюгів. Реплікація відбувається в ядрі в 8-період інтерфази. На цей час хромосоми під світловим мікроскопом не виявляються.
Реплікація ДНК — найважливіший молекулярний процес, що лежить в основі всіх різновидів поділу клітин, усіх типів розмноження, а отже, в основі забезпечення тривалого існування окремих індивідів, популяцій і всіх видів живих організмів. Для кожного виду дуже важливо підтримувати сталість свого генотипу, тобто зберігати незмінність нуклеотидної послідовності генотипу. Для цього необхідно абсолютно точно відтворювати молекули ДНК перед кожним поділом клітини, тобто основне функціональне значення реплікації — забезпечити нащадка стабільною генетичною інформацією розвитку, функціонування і поведінки.
Напівконсервативний шлях реплікації ДНК. Установлено (М. Мезельсон, Ф. Сталь), що в процесі реплікації дві нитки ДНК розділяються. Кожна з них є шаблоном (матрицею) для синтезу
вздовж неї нової нитки. Послідовність основ, які мають бути в нових нитках, можна легко передбачити, тому що вони комплементарні основам, наявним у старих нитках. Таким чином, утворюються дві дочірні молекули, ідентичні материнській. Кожна дочірня молекула складається з однієї старої (материнської) та однієї нової нитки. Оскільки тільки одна материнська нитка збережена в кожній дочірній молекулі, то такий тип реплікації має назву напівконсервативного.
Кожен з двох ланцюгів материнської молекули ДНК використовується як матриця для синтезу нових комплементарних ланцюгів.
Механізм реплікації ДНК. Реплікація ДНК — складний, багатоступінчастий процес, що потребує залучення великої кількості спеціальних білків і ферментів. Наприклад, ініціаторні білки утворюють реплікаційну вилку, ДНК-топоізомерази розкручують ланцюги, ДНК-гелікази і дестабілізаційний білок розщеплюють ДНК на два окремі ланцюги, ДНК-полімераза і ДНК-приймаза каталізують полімеризацію нуклеотидтрифосфатів і утворення нового ланцюга, ДНК-лігази руйнують РНК-затравки на ланцюгах ДНК, що відстають, та ін. Процес відбувається аналогічно як у прокаріотів, так і в еукаріотів, хоча дещо відрізняється за швидкістю, спрямованістю, кількістю точок реплікації тощо. Швидкість реплікації в еукаріотів дуже велика — 50 нуклеотидів за 1 с, а в прокаріотів ще вища — до 2000 нуклеотидів за 1 с.
Точність реплікації забезпечується комплементарною взаємодією азотистих основ матричного ланцюга і ланцюга, що будується. Крім цього, весь процес контролюється ДНК-полімеразою, що самостійно коригує та усуває помилки синтезу.
Основні етапи реплікації
1. Ініціація (від лат. іпіtialis — первинний, початковий). Це активація дезоксирибонуклеотидів. Монофосфати дезоксири-бонуклеотидів (АМФ, ГМФ, ЦМФ, ТМФ) перебувають у стані "вільного плавання" в ядрі і є "сировиною" для синтезу ДНК. Для включення в ДНК вони активуються в результаті взаємодії з АТФ. Ця реакція називається фосфорилюванням і каталізується ферментом фосфорилазою. При цьому утворюються трифосфати дезоксирибонуклеотидів, такі як АТФ, ГТФ, ЦТФ, ТТФ. У такому вигляді вони багаті на енергію й здатні до полімеризації.
Розпізнавання точки ініціації. ДНК починає розкручуватися з певної точки. Така особлива точка називається точкою ініціації реплікації (спеціальна послідовність нуклеотидів). Щоб визначити цю точку, потрібні специфічні білки-ініціатори. У вірусів і прокаріотів є тільки одна точка ініціації, а в еукаріотів, у яких вені кі молекули ДНК, може бути багато точок ініціації реплікації, що зрештою зливаються одна з одною при повному роз'єднанні ланцюгів ДНК. Реплікація обох ланцюгів ДНК відбувається одночасно і беззупинно.
Подвійна спіраль ДНК розкручується і розгортається на окре-мі нитки ДНК шляхом розриву слабких водневих зв'язків між комплементарними нуклеотидами. Цей процес забезпечують ферменти — гелікази. Оголені основи А, Т, Г і Ц обох ланцюгів проектуються в каріоплазму.
Ферменти, які називаються топоізомеразами, розривають і заново зшивають окремі нитки ДНК, допомагають розкручуванню спіралі. Завдяки роз'єднанню ланцюгів ДНК виникають репліка-цінні вилки. Нові нитки ДНК утворюються на кожному зі звіль-нених ланцюгів. їх ріст відбувається в протилежних напрямках.
2. Елонгація. Вільні трифосфати дезоксирибонуклеотидів своїми азотистими основами приєднуються водневими зв'язками до азотистих основ обох ланцюгів ДНК, відповідно до правила комп-лементарності, тобто А—Т, Ц—Г.
Елонгація — це додавання дезоксирибонуклеотиду до 3 -кінця ланцюга, що росте. Процес каталізується ДНК-полімеразою.
Трифосфати дезоксирибонуклеотидів (тринуклеотиди), приєднуючись до кожного ланцюга ДНК, розривають свої внутрішні високоенергетичні зв'язки й утворюють монофосфати дезоксирибонуклеотидів (мононуклеотиди), що є звичайними компонентами ДНК. При цьому в нуклеоплазму надходять пірофосфатні молекули, що звільнилися (Р—Р).
Утворення нових ланцюгів ДНК. Воно здійснюється в подальшому приєднанні сусідніх нуклеотидів, які зв'язуються між собою фосфорними залишками та утворюють новий ланцюг ДНК. Процес каталізується ферментом ДНК-полімеразою. При цьому необхідна наявність йонів металів Мп2+ або М£2\ ДНК-полімераза може полімеризувати дезоксирибонуклеотиди в напрямку 5'—З', тобто від вуглецевого З'-кінця молекул ДНК. Оскільки дві нитки ДНК антипаралельні, то нові нитки мають утворюватися на старих (материнських) у протилежних напрямках. Одна нова нитка виникає в напрямку 5'—3'. Ця нитка називається провідною. На другій материнській нитці формуються короткі сегменти ДНК у напрямку 3'—5'. Згодом вони об'єднуються і утворюють довгу відстаючу нитку.
Утворення праймерів. На відстаючій нитці спочатку формується короткий ланцюг РНК за шаблоном ДНК. Вона називається РНК-праймером і містить послідовність із 10—60 нуклеоти-дів. Фермент праймаза каталізує полімеризацію блоків РНК (А, У, Г, Ц) у праймері. РНК-праймер утворюється тому, що ДНК-полімераза не може ініціювати синтезу нової нитки ДНК у відстаючому ланцюгу в напрямку 3'—5', вона тільки може каталізувати її ріст. Праймери пізніше видаляються, а порожнини, які утворилися, заповнюються дезоксирибонуклеотидами ДНК у напрямку 5'—3', що завершує побудову другого ланцюга. На місці праймерів утворюються фрагменти нового ланцюга ДНК, які називаються фрагментами Оказакі; вони складаються зі 100—200 нуклеотидів. Ці фрагменти зшиваються полінуклеотидлігазами, у результаті чого утворюється другий повноцінний ланцюг. Цей процес називається дозріванням.
Редагування. Завдяки чіткій комплементарності пар основ редагування забезпечує точну реплікацію ДНК. Проте іноді виникають помилки в приєднанні основ, вони видаляються ДНК-полімеразою, яка для цього знову зв'язується з молекулами ДНК (репарація).
3. Термінація (від лат. terminaton — кінцевий). Коли завершився процес реплікації, молекули, що утворилися, розділяються, і кожна дочірня нитка ДНК скручується разом із материнською в подвійну спіраль. Завдяки цьому утворюються дві молекули ДНК, ідентичні материнській. Вони формуються окремими фрагментами по довжині хромосоми. Такий окремий фрагмент ДНК, що подвоюється на одній хромосомі, називається репліконом. Виникає відразу декілька репліконів, причому асинхронно й у різних її ділянках. Процес реплікації стосується всієї хромосоми й перебігає практично одночасно, з однаковою швидкістю. Після завершення реплікації в репліконах вони зшиваються ферментами в одну молекулу ДНК. У людській клітині, що ділиться, утворюється понад 50 000 репліконів одночасно. Довжина кожного з них — 30 мкм. Завдяки великій кількості репліконів швидкість реплікації збільшується в тисячі разів. Процес подвоєння генетичного матеріалу триває майже 10 год. Ділянки хромосом, де починається реплікація, називаються точками ініціації. Вважають, що це, ймовірно, місця прикріплення інтерфазних хромосом до
Розділ 2
57
кілків ламели ядерної оболонки. Процес включається цитоплазматичним фактором невідомої природи, що надходить у ядро.
Реплікація перебігає в строго визначеному порядку, тобто спо-• 11 ггку починають реплікуватись одні ділянки хромосом, а пізніше — інші. У синтетичному періоді інтерфази подвоюється також і кількість гістонових білків, що асоціюються із синтезованими ДНК і утворюють класичну структуру хроматину. Порушення точності реплікації призводить до порушення синтезу білків і розвитку патологічних змін клітин і органів.
Реплікація має велике значення, а саме:
є важливим молекулярним механізмом, який лежить в основі всіх різновидів поділу клітин прокаріотів та еукаріо-тів;
забезпечує всі типи розмноження одноклітинних та багатоклітинних організмів;
підтримує сталість клітинного складу органів, тканин і організму внаслідок фізіологічної регенерації;
забезпечує тривале існування окремих індивідів;
забезпечує тривале існування видів організмів;
процес сприяє точному подвоєнню інформації;
у процесі реплікації можливі помилки (мутації), що може призводити до порушень синтезу білків з розвитком патологічних змін.