Протисты
Протистами (Protista) называют организмы, тело которых состоит из одной-единственной эукариотической клетки. Другой термин, употребляемый для обозначения одноклеточных существ – простейшие, – постепенно выходит из употребления, потому что соответствующее ему латинское название Protozoa традиционно обозначает тип царства животных. А провести четкую границу между протистами-животными и протистами-водорослями практически невозможно – ряд из них могут использовать как автотрофный (фотосинтез), так и гетеротрофный (т.е. потреблять готовые органические вещества) типы питания. Поэтому все больше ученых соглашается с тем, что эти организмы следует выделить в отдельное царство.
Размеры протистов – обычные размеры эукариотических клеток, примерно 0,02–0,2 мм, но «в их лице» мы встречаемся с самой сложной организацией, известной для биологических клеток вообще. Ведь каждый протист – самостоятельный, и в его клетке-теле происходят такие процессы, которые у многоклеточных осуществляются лишь на уровне тканей и органов.
Как все эукариотические клетки, протисты окружены клеточной оболочкой – плазматической мембраной (плазмалеммой), к которой снаружи примыкает мукоидный (т.е. содержащий остатки молекул углеводов) слой – гликокаликс. В этом слое у протистов присутствуют молекулы, воспринимающие прикосновения, различные запахи, свет и т.д., т.е. играющие роль органов чувств у многоклеточных. Снаружи клеточной оболочки у многих протистов могут располагаться различные типы дополнительных оболочек, органические или минеральные (кремнеземные) чешуйки, раковинки, а у протистов-водорослей – целлюлозная стенка.
Клетки протистов содержат весь комплекс органелл, характерный для клеток эукариотических организмов: это и эндоплазматическая сеть, и аппарат Гольджи, и митохондрии, и пластиды. Однако в цитоплазме протистов присутствуют еще и дополнительные органеллы, необходимые им как самостоятельным организмам, – сократительные вакуоли и экструсомы, или стрекательные органеллы. Сократительные вакуоли поддерживают необходимую концентрацию воды в клетке. По мере накопления избытка жидкости они либо сливаются с клеточной оболочкой, после чего их содержимое оказывается в окружающей среде, либо (у инфузорий) сокращаются, «выдавливая» лишнюю воду через особую пору в клеточной оболочке. Стрекательные органеллы протистов также способны выбрасывать свое содержимое наружу – в ответ на внешнее раздражение. Ядовитые, жгучие, парализующие вещества используются для нападения и защиты, а у некоторых протистов-паразитов (например, у микро- и книдоспоридий) – для проникновения в клетку хозяина
Очень важные органеллы, присутствующие в клетках протистов, – жгутики (у жгутиконосцев) и реснички (у инфузорий). Строение их одинаковое, но характер движения различен, а кроме того, реснички короче жгутиков. И те, и другие могут использоваться как для передвижения, так и для захвата пищи.
Клетки некоторых протистов содержат не одно, а несколько, иногда даже очень много, ядер. Относительно крупные клетки, в цитоплазме которых присутствуют многочисленные однотипные ядра, называют плазмодиями. А фораминиферам и инфузориям присущ так называемый ядерный дуализм: они обладают двумя типами ядер – вегетативным (макронуклеусом) и генеративным (микронуклеусом). Вегетативное ядро может содержать множественный набор хромосом (быть полиплоидным) и обеспечивает повседневную жизнь клетки: в нем образуются молекулы РНК, необходимые для синтеза белка. Генеративное же ядро в этих процессах участия не принимает, содержит «нормальный» (двойной или одинарный) набор хромосом и активизируется только во время деления клетки.
Существуют и протисты, тело которых состоит не из одной, а из нескольких или многих клеток. Однако такие «тела» представляют собой не единые организмы, а колонии одноклеточных существ, поскольку каждая клетка в них остается относительно независимой.
Размножение протистов может происходить разными способами. Наиболее простой – обычное деление клетки путем митоза, в результате чего из одной особи возникают две новых. Такое бесполое размножение наблюдается у всех протистов, вне зависимости от того, содержат ли их ядра одинарный (гаплоидный) или двойной (диплоидный) набор хромосом. Но у многих групп этих существ жизненный цикл включает чередование диплоидного и гаплоидного поколений. В этом случае стадии с одинарным набором хромосом – по аналогии с половыми клетками многоклеточных животных и растений – считаются гаметами. Ядра таких клеток-гамет способны сливаться, и в результате образуется клетка с двойным набором хромосом. В этом случае говорят о наличии у данного вида полового процесса, т.е. рекомбинации генетического материала разных особей, в то время как при обычном делении дочерние клетки генетически идентичны родительской. В образовавшейся диплоидной клетке в свое время происходит редукционное деление – мейоз, вновь приводящее к образованию гаплоидных гамет, участвующих в следующем половом цикле. Однако до слияния гаплоидные клетки многих протист могут многократно делиться и жить совершенно самостоятельно. У ряда видов преобладающей в жизненном цикле является именно гаплоидная стадия, а для некоторых неизвестно ни их слияние, ни диплоидные ядра, ни, соответственно, половой процесс. У инфузорий, наоборот, существует половой процесс, но не образуется гамет: клетки-партнеры просто обмениваются образовавшимися в ходе мейоза гаплоидными ядрами. В этом случае говорят, что половой процесс не связан с процессом размножения, ведь число инфузорий при этом не увеличивается.
У многих, особенно паразитических, протистов в ходе жизненного цикла происходит сложная смена стадий, различающихся не только числом хромосом в ядре, но и формой, и строением клетки. Таких стадий может быть довольно много, и каждая из них предназначена для выполнения определенных функций – обитания вне и внутри клетки организма-хозяина, в одном или другом хозяине, для многократного деления или для расселения.
Распространены протисты на земном шаре повсеместно. Основная среда их обитания – вода. Причина этого проста: в активном состоянии клетки этих организмов не способны противостоять высыханию. Зато встречаются простейшие практически во всех водоемах, от луж до океанов. Много этих организмов обитает и в почве, населяя тонкую пленку воды, обволакивающую ее частицы.
Еще одна среда обитания протистов – организмы животных и растений, где они могут жить как в качестве безразличных для хозяина сожителей-комменсалов (т.е. безразличных сожителей), так и опасных паразитов. Целый ряд видов простейших – возбудители опасных заболеваний животных и человека, например малярии, сонной болезни, пендинской язвы... С другой стороны, ряд протистов вступает с организмом хозяина во взаимовыгодные симбиотические отношения. Например, простейшие, обитающие в пищеварительном тракте растительноядных животных, помогают им усваивать энергию, содержащуюся в плохо перевариваемой животным организмом целлюлюзе, и синтезируют незаменимые аминокислоты.
Тропическая лейшмания вызывает пендинскую язву |
Жизненный цикл гамбийской трипаносомы |
Попадая в неблагоприятную для себя среду, многие из протистов не гибнут, а образуют неактивные стадии – цистыи споры, покрытые плотными оболочками. В таком состоянии они способны выдерживать широкий диапазон внешних условий, в том числе и полное высыхание, а при попадании в благоприятную среду вновь «оживают». У некоторых видов протистов споры всегда образуются при наступлении неблагоприятных условий, у других – к спорообразованию способны только отдельные стадии, возникающие, например, осенью. Споры, переносимые водой, ветром, животными, – основной способ расселения тех протистов, чья активная жизнь проходит в других организмах.
Питаться протисты могут как путем экзоцитоза, т.е. всасывая мельчайшие пищевые частицы и молекулы органических веществ через оболочку клетки, так и хищничая – активно захватывая и переваривая, например, бактерий или своих соплеменников. Представители некоторых видов, как уже было сказано, имеют хлоропласты и могут получать питательные вещества в процессе фотосинтеза. В сообществах такие фотосинтезирующие формы выполняют роль продуцентов. Особенно важна эта роль в водоемах, где на их долю может приходиться до 80% первичной продукции, т.е. синтезируемого органического вещества. Гетеротрофные же протисты играют роль редуцентов, вновь включающих в пищевые цепи мертвые органические остатки, но могут быть и хищниками первого звена. Сами они, как и фотосинтезирующие формы – одноклеточные водоросли, – служат пищей многочисленным мелким беспозвоночным и малькам рыб, являясь первым (или вторым) звеном пищевых цепей в почвенных и водных сообществах.
Что касается систематики протистов, то здесь у специалистов нет единого мнения. Чаще всего принимается предложенное еще в 1882 г. и использованное в этой статье деление всего многообразия одноклеточных на шесть групп – саркодовых (Sarcodina), жгутиконосцев (Mastigophora), инфузорий (Ciliophora), споровиков (Sporozoa), микроспоридий (Microspora) и книдоспоридий (Cnidospora). При рассмотрении протистов как отдельного царства этим группам придается, соответственно, ранг типов. В противном случае это могут быть классы в типе простейших царства животных, в то время как ботаники относят «своих» протистов к разным отделам царств растений и грибов.
Очевидно, однако, что далекими предками всех населяющих Землю многоклеточных животных, растений и грибов были одноклеточные гетеротрофные организмы. Причем разные их группы дали начало разным таксонам. Например, эвгленовые водоросли, вероятно, произошли от жгутиконосцев класса кинетопластид – об этом свидетельствует ряд тонких деталей строения их клетки. А представители класса воротничковых жгутиконосцев, похоже, дали начало животным. Во всяком случае, строение их клеток удивительно похоже на строение воротничковых клеток губок. Воротничковые же жгутиконосцы, как показывают исследования молекулярных биологов, вероятно, были предками и настоящих грибов.
Как бы там ни было, о протистах, клетки которых несут «растительные» и «грибные» признаки, на страницах «Биологии» уже рассказывалось* . Поэтому данная статья будет посвящена в первую очередь тем одноклеточном организмам, которых традиционно принято рассматривать при изучении курса зоологии.
Размеры протистов – обычные размеры эукариотических клеток, примерно 0,02–0,2 мм, но «в их лице» мы встречаемся с самой сложной организацией, известной для биологических клеток вообще. Ведь каждый протист – самостоятельный, и в его клетке-теле происходят такие процессы, которые у многоклеточных осуществляются лишь на уровне тканей и органов.
Р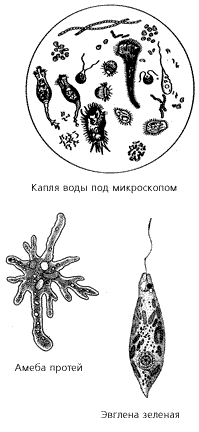 азнообразные
протисты – инфузории, амебы, солнечники
– присутствуют в любом водоеме, в любой
луже. Даже в обычном комнатном аквариуме
или горшке с комнатным растением можно
обнаружить подчас совершенно фантастических
созданий. Однако невооруженным глазом
их не увидишь. Нужен микроскоп или
бинокулярная лупа с увеличением в 20–100
раз. Еще нам потребуется несколько чашек
Петри или предметных стекол, обычные
пипетки и иглы.
азнообразные
протисты – инфузории, амебы, солнечники
– присутствуют в любом водоеме, в любой
луже. Даже в обычном комнатном аквариуме
или горшке с комнатным растением можно
обнаружить подчас совершенно фантастических
созданий. Однако невооруженным глазом
их не увидишь. Нужен микроскоп или
бинокулярная лупа с увеличением в 20–100
раз. Еще нам потребуется несколько чашек
Петри или предметных стекол, обычные
пипетки и иглы.
В естественных водоемах пробы для изучения простейших лучше собирать газовым сачком, проводя им по зарослям водных растений и придонному слою воды. Если у вас нет сачка, в пруду можно зачерпнуть кусочек грунта, придонной взвеси, тины, бахрому с листьев водных растений или коряг. Весь собранный материал немедленно помещают в небольшую банку с водой.
Если в вашем распоряжении имеется оптический прибор с увеличением в 20–50 раз, тщательно перемешайте собранный материал и перелейте его в чашку Петри так, чтобы толщина слоя воды не превышала 0,5 см. При работе с микроскопом с большим увеличением на дно чашки переносится только капля воды с материалом. Можно поместить каплю и на предметное стекло, а чтобы она не высохла сразу, закрыть ее покровным стеклом. Для того чтобы оно своей тяжестью не раздавило всю живность, по четырем краям покровного стекла нужно сделать небольшие «ножки» из пластилина.
Отбирая пробу, не жадничайте: рассматривать ее надо в проходящем освещении, т.е. когда свет от зеркала попадает в объектив микроскопа через образец, и если непрозрачные частицы будут лежать на стекле сплошным слоем, вы просто ничего не увидите. При небольшом увеличении можно рассматривать относительно толстую каплю и «путешествовать» не только в стороны, но и в глубь этого маленького мира, поднимая или опуская объектив.
Протисты со жгутиками
Протистов, имеющих жгутики, относят к типу жгутиконосцев (Mastigophora). Именно в пределах этой группы помимо организмов, питающихся исключительно готовыми органическими веществами, встречаются формы, обладающие хлоропластами и способные к фотосинтезу. Многие же из гетеротрофных жгутиконосцев являются паразитами, возбудителями опасных заболеваний. Хотя деление жгутиконосцев на «животных» и «растительных» условно, в этой статье, как уже было сказано, мы остановимся на описании только гетеротрофных представителей этого типа.
Класс жгутиконосцев-кинетопластид (Kinetoplastea) включает около 150 видов. Характерной чертой клеток этих организмов является единственная крупная митохондрия, простирающаяся на всю длину клетки в виде разветвленной сети. В районе основания жгутика в этой митохондрии имеется обширная область – кинетопласт, в которой сосредоточена митохондриальная ДНК. Ядра кинетопластид несут одинарный набор хромосом, а размножаются эти протисты простым делением на двое. Половой процесс у них неизвестен.
М уха
цеце
уха
цеце
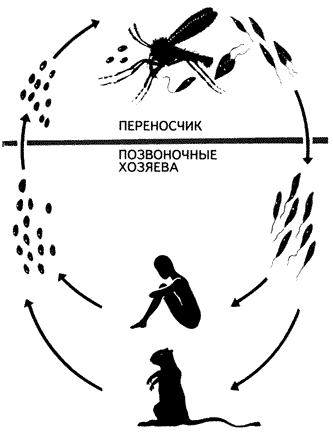
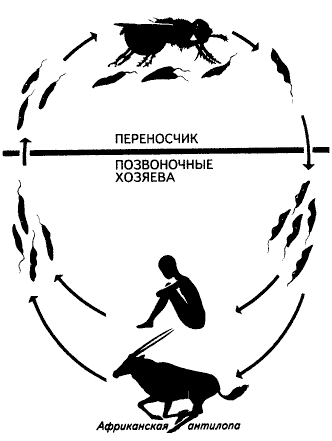
К группе одножгутиковых кинетопластид,или трипаносоматид(Trypanosomida) относится около 100 видов простейших. Все они – паразиты многоклеточных организмов. Некоторые (низшие формы) проводят всю жизнь в организме одного хозяина – какого-нибудь беспозвоночного животного. Другие (их считают более высокоорганизованными) на протяжении жизненного цикла меняют хозяев, переселяясь из беспозвоночного (обычно насекомого) в организм позвоночного животного, а в отдельных случаях – и растения (жгутиконосцы из рода Phytomonas). Беспозвоночным хозяином возбудителя сонной болезни – трипаносомыTrypanosoma brucei является муха це-це. При укусе с ее слюной в кровь человека или животного попадают подвижные жгутиковые стадии паразита, дальнейшее развитие которых, состоящее в ряде последовательных делений, происходит в кровяном русле. Кровь разносит трипаносом по организму, и они переходят в клетки тканей, где образуют формы, лишенные жгутиков. После разрушения клеток паразиты вновь оказываются в крови, а оттуда могут попасть и в кишечник новой мухи це-це. В Южной Америке сонной болезни аналогична болезнь Чагаса, вызываемая трипаносомой T.cruzi и переносимая триатомовыми (поцелуйными) клопами.
У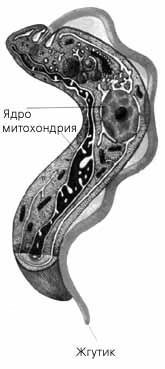 некоторых видов трипаносом (род Trypanosoma)
более древняя стадия, связанная с
беспозвоночным хозяином, может вторично
выпадать, как, например, у T.equiperdum –
возбудителя случной болезни лошадей,
передающейся у этих копытных половым
путем.
Представители
рода Leishmania из
группы трипаносомид – возбудители
тяжелых заболеваний человека и животных
– лейшманиозов.
Различают кожный лейшманиоз, или
пендинскую язву (возбудители
– L.major и L.tropica),
кожно-слизистый (L.braziliensis)
и висцеральный (L.donovani)
лейшманиозы. Беспозвоночным хозяином
лейшманий являются москиты. Питаясь на
позвоночном животном, они вводят со
слюной в его ткани подвижные жгутиковые
клетки паразита, которые в межклеточных
пространствах захватываются
лейкоцитами-макрофагами, но не
перевариваются в них, а превращаются в
безжгутиковые формы, переходящие к
внутриклеточному паразитированию и
делению. Пораженная клетка со временем
лопается, и паразиты переходят в соседние
клетки и ткани. Вновь попадая с кровью
в пищеварительный тракт москита, они
опять превращаются в подвижные жгутиковые
формы. Лейшманиозы встречались в
среднеазиатских республиках бывшего
СССР.
некоторых видов трипаносом (род Trypanosoma)
более древняя стадия, связанная с
беспозвоночным хозяином, может вторично
выпадать, как, например, у T.equiperdum –
возбудителя случной болезни лошадей,
передающейся у этих копытных половым
путем.
Представители
рода Leishmania из
группы трипаносомид – возбудители
тяжелых заболеваний человека и животных
– лейшманиозов.
Различают кожный лейшманиоз, или
пендинскую язву (возбудители
– L.major и L.tropica),
кожно-слизистый (L.braziliensis)
и висцеральный (L.donovani)
лейшманиозы. Беспозвоночным хозяином
лейшманий являются москиты. Питаясь на
позвоночном животном, они вводят со
слюной в его ткани подвижные жгутиковые
клетки паразита, которые в межклеточных
пространствах захватываются
лейкоцитами-макрофагами, но не
перевариваются в них, а превращаются в
безжгутиковые формы, переходящие к
внутриклеточному паразитированию и
делению. Пораженная клетка со временем
лопается, и паразиты переходят в соседние
клетки и ткани. Вновь попадая с кровью
в пищеварительный тракт москита, они
опять превращаются в подвижные жгутиковые
формы. Лейшманиозы встречались в
среднеазиатских республиках бывшего
СССР.
Общий вид и строение трипаносомы
Группа двужгутиковых кинетопластид (Bodonida) включает около 50 видов свободно живущих жгутиконосцев. Они обитают в лужах и прудах и ведут хищный образ жизни: приближаясь к своим одноклеточным жертвам, парализуют их находящимися вблизи оснований жгутиков стрекательными органеллами, а после этого заглатывают при помощи специализированного ротового аппарата.
Класс жгутиконосцев-дипломонад (Diplomonadea) включает около 40 видов протист, в клетках которых отсутствуют такие важные органеллы, как митохондрии и аппарат Гольджи. Считается, что эти существа – наиболее примитивные из всех ныне живущих эукариотических организмов. Дипломонады – паразиты пищеварительного тракта различных, как беспозвоночных, так и позвоночных, животных. Обычно они имеют 8 жгутиков, один из которых обращен назад и втянут в особую ложбинку – «клеточный рот», через который дипломонады, как и многие другие паразитические жгутиконосцы, всасывают из окружающей среды питательные вещества. К дипломонадам относятся лямблии (Lamblia) – повсеместно распространенные паразиты желудочно-кишечного тракта млекопитающих, в том числе и человека. Эти жгутиконосцы обладают брюшным присасывательным диском, с помощью которого присасываются к поверхности кишечника. Размножаясь, лямблии попросту блокируют всасывание питательных веществ через стенки кишечника, вызывая кровавые поносы – лямблиозную диарею. Попадая в задний отдел кишечника, эти паразитические протисты втягивают жгутики и покрываются толстой оболочкой, превращаясь в расселительную стадию – цисту, которая выносится в окружающую среду и может заразить нового хозяина. Заражение происходит при употреблении сырой воды и загрязненных продуктов питания.
Класс опалин (Opalinea) включает около 50 видов крупных – до нескольких миллиметров в длину – протистов, живущих в качестве комменсалов в кишечнике земноводных. Клетки опалин содержат много одинаковых ядер, а снаружи покрыты многочисленными, расположенными рядами жгутиками. Клеточного рта у них нет, и питательные вещества они поглощают всей поверхностью тела. У опалин известны как бесполое размножение (деление надвое, при котором многочисленные ядра распределяются между делящимися клетками, а потом восстанавливаются в числе за счет митотических делений), так и половой процесс.
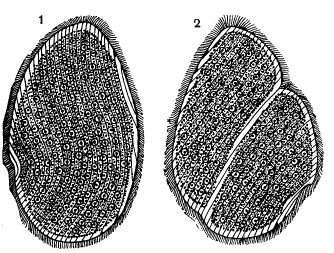
Паразитический жгутиконосец из задней кишки лягушки (Opalina ranarum). Видно большое количество жгутиков и ядер. 1 – неделящаяся особь; 2 – деление, борозда проходит косо по отношению к длинной оси простейшего
Последний происходит в кишечнике головастиков, заглатывающих цисты опалин, выходящие в воду при размножении лягушек. В клетках, появившихся из цист, происходит мейоз, и образуются одноядерные макро- и микрогаметы. Они сливаются, и возникает диплоидная зигота, из которой развивается взрослая опалина.
Биология опалин хорошо иллюстрирует возможность удивительно тонкого приспособления жизненного цикла паразитов (в данном случае, правда, безвредных) к ситуации, когда представители вида-хозяина размножаются или встречаются между собой нерегулярно. Опалины образуют цисты и выходят наружу раз в году, весной, когда лягушки приступают к икрометанию. Там они (цисты) живут некоторое время в ожидании, не заглотит ли их вышедший из лягушачьей икринки головастик. Разумеется, шанс невелик, но другого пути попасть в организм нового хозяина у опалин нет. Если головастик заглотит цисту, то в вышедшей из нее клетке происходит несколько делений, одно из которых – редукционное. В результате образуются гаплоидные клетки-гаметы, которые тут же, в кишечнике, сливаются и дают начало новым опалинам, которые будут очень долго жить в кишечнике лягушки, не образуя цист, – до тех пор, пока земноводное не повзрослеет и не приступит к размножению. Можно предположить, что стимулом к образованию расселительных цист у этих жгутиконосцев служит изменение гормонального фона в крови хозяина.
Класс парабазалиевых жгутиконосцев (Parabasalea) включает около 100 видов паразитических простейших, клетки которых лишены митохондрий, но, как правило, содержат несколько ядер и отличаются крупными размерами – до 1 мм в длину. Представители более примитивного отряда – трихомонад (Trichomonadida) имеют 4 жгутика, а у более высокоорганизованных гипермастигид (Hypermastigida) их количество может достигать нескольких тысяч.
Некоторые трихомонады паразитируют в кишечнике и мочеполовом тракте млекопитающих и птиц, вызывая заболевания соответствующих систем органов (заражение осуществляется преимущественно через внешнюю среду). Большинство же представителей класса парабазалиевых – симбионты кишечника древесиноядных термитов и тараканов, питающиеся поступающей в кишечник хозяев пищей и обитающими здесь бактериями.
Симбиоз термитов и обитающих в их кишечнике жгутиконосцев, а также азотфиксирующих бактерий и бактерий, перерабатывающих целлюлозу – еще один пример совершенного приспособления живых организмов к окружающей среде. Ведь ряд видов термитов питается почти исключительно мертвой древесиной, представляющей из себя фактически чистую целлюлозу – продукт, содержащий значительное количество энергии, но практически непереваримый в организме животных. Необходимые ферменты имеются в достаточном количестве лишь у представителей мира одноклеточных. Именно их, своих постояльцев (или «домашних животных»?) термит и «кормит» древесиной. Микроорганизмы, способные переваривать целлюлозу, в свою очередь, делятся полученной энергией с бактериями, способными химически связывать свободный азот – ведь в мертвой древесине практически не остается белка. В итоге сожители кишечника термита накапливают в своих клетках питательные вещества, вполне доступные для переваривания самому термиту и содержащие не только энергию, но и белок, включающий все необходимые насекомому виды аминокислот.
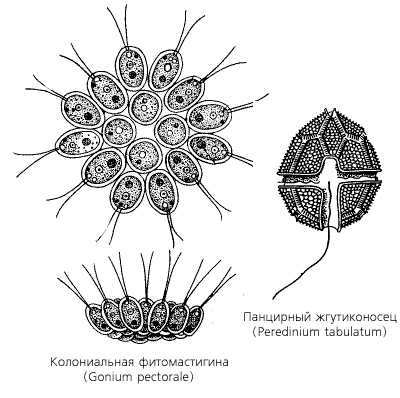
Класс воротничковых жгутиконосцев (Choanoflagellatеa) включает 100 видов мелких (0,005 – 0,02 мм) организмов, клетки которых имеют один жгутик. Основание этого жгутика окружено венчиком микроворсинок, называемым воротничком и служащим для отфильтровывания взвешенных в воде пищевых частиц (бактерий), подгоняемых током воды к основанию жгутика. С внешней стороны вблизи основания воротничка образуются мелкие ложноножки (псевдоподии), захватывающие из воды взвесь питательных веществ. Воротничковые – свободноживущие протисты, среди которых имеются как планктонные (т.е. свободно плавающие), так и сидячие; как одиночные, так или колониальные формы. Ядра воротничковых жгутиконосцев содержат двойной набор хромосом, но половой процесс у них неизвестен.
Рассмотреть в микроскоп в домашних условиях паразитических жгутиконосцев по понятным причинам не так-то просто. Зато достаточно легко познакомится с зелеными и эвгленовыми водорослями, которых легко опознать по содержащемся в теле хлоропластам. Рассматривая материал, собранный летом в пруду, в первую очередь находишь как раз окрашенные хлорофиллом автотрофные организмы.
Отличить зеленые водоросли от эвгленовых можно по наличию двух одинаковых передних жгутиков, тогда как у эвглен второй жгутик недоразвит. Клетка эвглены имеет веретенообразную форму и продвигается достаточно быстро за счет движения жгутика. Находящиеся под плазмалеммой тела эвглены микротрубочки способствуют совершению ею «червеобразных», извивающихся движений.
Эвглену, а также колониального жгутиконосца вольвокс (Volvox), относимого ботаниками к отделу зеленых водорослей, легко найти в середине лета и осенью в зачерпнутой со дна пруда воде или поводив сачком по растущим вблизи дна водным растениям. Другие зеленые водоросли, в том числе хламидомонаду (Chlamydomonas) проще всего увидеть в «цветущей» – зеленоватого цвета – летней воде небольших, но постоянных луж и прудиков.
Класс парабазалиевых жгутиконосцев (Parabasalea) включает около 100 видов паразитических простейших, клетки которых лишены митохондрий, но, как правило, содержат несколько ядер и отличаются крупными размерами – до 1 мм в длину. Представители более примитивного отряда –трихомонад (Trichomonadida) имеют 4 жгутика, а у более высокоорганизованных гипермастигид(Hypermastigida) их количество может достигать нескольких тысяч.
Некоторые трихомонады паразитируют в кишечнике и мочеполовом тракте млекопитающих и птиц, вызывая заболевания соответствующих систем органов (заражение осуществляется преимущественно через внешнюю среду). Большинство же представителей класса парабазалиевых – симбионты кишечника древесиноядных термитов и тараканов, питающихся поступающей в кишечник хозяев пищей и обитающими здесь бактериями.
Симбиоз термитов и обитающих в их кишечнике жгутиконосцев, а также азотфиксирующих бактерий и бактерий, перерабатывающих целлюлозу, – еще один пример совершенства приспособления живых организмов к окружающей среде. Ведь ряд видов термитов питается почти исключительно мертвой древесиной, представляющей из себя фактически чистую целлюлозу – продукт, содержащий значительное количество энергии, но практически неперевариваемый в организме животных. Необходимые ферменты имеются в достаточном количестве лишь у представителей мира одноклеточных. Именно их, своих постояльцев (или «домашних животных»), термит и «кормит» древесиной. Микроорганизмы, способные переваривать целлюлозу, в свою очередь, делятся полученной энергией с бактериями, способными химически связывать свободный азот – ведь в мертвой древесине практически не остается белка. В итоге сожители кишечника термита накапливают в своих клетках питательные вещества, вполне доступные для переваривания самому термиту и содержащие не только энергию, но и белок, включающий все необходимые насекомому виды аминокислот.

Различные жгутиконосцы из кишечника термитов: A – Teratomipha mirabilis; Б – Spirotrichonympha flagellata; B – Coronympha octonaria; Г – Calonympha grassi; Д – Trichonympha turkestana; E – Rhynchonympha tarda; 1 – ядро; 2 – аксостили
Класс воротничковых жгутиконосцев (Choanoflagellatеa) включает 100 видов мелких (0,005 – 0,02 мм) организмов, клетки которых имеют один жгутик. Основание этого жгутика окружено венчиком микроворсинок, называемым воротничком и служащим для отфильтровывания взвешенных в воде пищевых частиц (бактерий), подгоняемых током воды к основанию жгутика. С внешней стороны вблизи основания воротничка образуются мелкие ложноножки (псевдоподии), захватывающие из воды взвесь питательных веществ. Воротничковые – свободноживущие протисты, среди которых имеются как планктонные (т.е. свободноплавающие), так и сидячие; как одиночные, так и колониальные формы. Ядра воротничковых жгутиконосцев содержат двойной набор хромосом, но половой процесс у них неизвестен.
Рассмотреть в микроскоп в домашних условиях паразитических жгутиконосцев по понятным причинам не так-то просто. Зато достаточно легко познакомиться с зелеными и эвгленовыми водорослями, которых легко опо–знать по содержащимся в теле хлоропластам. Рассматривая материал, собранный летом в пруду, в первую очередь находишь как раз окрашенные хлорофиллом автотрофные организмы. Отличить зеленые водоросли от эвгленовых можно по наличию двух одинаковых передних жгутиков, тогда как у эвглен второй жгутик недоразвит. Клетка эвглены имеет веретенообразную форму и продвигается достаточно быстро за счет движения жгутика. Находящиеся под плазмалеммой тела эвглены микротрубочки способствуют совершению ею «червеобразных», извивающихся движений. Эвглену, а также колониального жгутиконосца вольвокс (Volvox), относимого ботаниками к отделу зеленых водорослей, легко найти в середине лета и осенью в зачерпнутой со дна пруда воде или поводив сачком по растущим вблизи дна водным растениям. Другие зеленые водоросли, в том числе хламидомонаду (Chlamydomonas), проще всего увидеть в «цветущей» – зеленоватого цвета – летней воде небольших, но постоянных луж и прудиков.
Саркодовые
К типу саркодовых (Sarcodina) относят протистов, способных образовывать так называемые ложноножки, или псевдоподии, – подвижные выросты цитоплазмы, выдающиеся за общие контуры тела клетки. Ложноножки саркодовых могут быть лопастевидными или цилиндрическими, нитевидными, ветвящимися и сливающимися между собой наподобие сетки. Бывает, что они имеют опорный каркас из продольных микротрубочек. Форма и строение ложноножек служат тем признаком, на основе которого саркодовых разделяют на отдельные классы и отряды. Большинство саркодовых – свободноживущие хищные организмы, питающиеся одноклеточными водорослями, жгутиконосцами, инфузориями, а также бактериями, которых они захватывают своими ложноножками и переваривают. Распространены саркодовые по всему земному шару и встречаются в водоемах с различной соленостью, а также в почве.
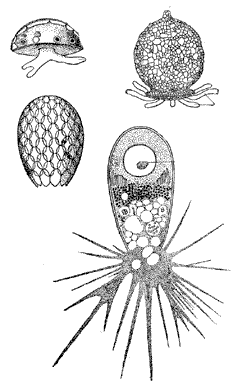
Различные виды раковинных амеб
Класс корненожек (Rhizopoda) включает несколько отрядов. К отряду настоящих амеб (Euamoebida) относится 200–250 видов протистов с лопастевидными псевдоподиями, с помощью которых они «ползают» по субстрату, и не имеющих каких-либо раковинок, свойственных другим корненожкам. Некоторые виды имеют веерную форму, с расширенным передним концом, на котором и образуются псевдоподии, другие – цилиндрическую и при активном движении образуют только одну переднюю псевдоподию. Размеры клеток этих организмов – от 0,005 до 0,02 мм.
Большинство настоящих амеб – донные организмы, обитающие в толще осадка. Однако иногда – для того, чтобы пересилиться на новое место, – они могут на непродолжительное время округляться и выпускать более длинные и тонкие (лучистые) псевдоподии, благодаря которым парят в толще воды и переносятся ее потоком. Размножаются настоящие амебы простым митотическим делением на–двое. Ядро клеток этих организмов содержит двойной набор хромосом, но до настоящего момента никто и никогда не наблюдал у них полового процесса.
Из представителя отряда настоящих амеб наиболее широко известна крупная (0,2–0,3 мм) пресноводная амеба-протей (Amoeba proteus). Именно о ней рассказывается в школьном учебнике зоологии. Представители другого рода, Acanthamoeba, обитают в почве. Однако некоторые виды акантамеб способны к факультативному (т.е. необязательному) паразитизму на теплокровных животных. Попадая в их организм с водой, они размножаются в межтканевой жидкости и могут вызывать у людей, страдающих иммунодефицитом, серьезные формы энцефалита.
Отряд корненожек шизопиренид (Schizopyrenida) включает около 100 видов мелких (0,005 – 0,01 мм) преимущественно почвенных протистов. От настоящих амеб их отличает присутствие на переднем конце пульсирующей зоны («гиалиновой шапочки»), а также способность большинства видов образовывать особые расселительные стадии, снабженные 2–4 жгутиками. Размножаются шизопирениды, как и настоящие амебы, простым митотическим делением надвое, половой процесс у них неизвестен.
Пресноводные амебы из этого отряда, относящиеся к роду Naegleria, при попадании в организм теплокровных животных способны, подобно акантамебам, образовывать временно паразитические формы, вызывающие менингоэнцефалит.
К отряду энтамеб (Entamoebida) относятся около 50 видов протистов, живущих в кишечном тракте позвоночных животных. Там они питаются как попадающей туда пищей, так и тканями самого кишечника, но обычно не наносят организму хозяина какого-либо существенного вреда. Однако энтамеба видаЕntamoeba histolitica, обитающая в кишечнике человека, в определенных условиях образует особую форму, проникающую в околокишечные ткани и печень и разрушающую их, а также поедающую эритроциты. Такое заболевание называется амебной дизентерией и встречается в тропических странах. Представители того же вида энтамеб, живущие в кишечнике жителей средней полосы, опасной формы не образуют.
Характерная особенность энтамеб – отсутствие в их клетках митохондрий и аппарата Гольджи. Однако, вероятно, это не примитивная черта, а вторичное упрощение – ведь в условиях кишечника митохондрии, отвечающие за кислородное дыхание, просто не нужны.
Отряд раковинных амеб (Testacida) включает около 300 видов простейших, тело которых окружено однокамерной раковинкой, в которой имеется устье для выхода псевдоподий. Раковинка эта может быть построена из белка, близкого по своему составу кератину, образующему наши волосы и ногти, из выделяемых клеткой кремнеземных пластинок или сцементированных песчинок. Обычный размер раковинки – 0,05–0,2 мм. Раковинные амебы встречаются преимущественно в пресных водоемах и в почве, а в морях, напротив, редки. Размножаются эти протисты митотическим делением надвое, причем одна из получившихся особей остается в старой раковинке, а другая окружает себя новой. Однако у раковинных амеб имеется и половой процесс, причем у разных форм он может протекать по-разному. В одних случаях ядра раковинных амеб несут двойной набор хромосом, но в определенный момент клетка образует цисту, в которой происходит редукционное деление. Возникает пара гаплоидных половых ядер, которые потом вновь сливаются друг с другом, – такой половой процесс называется автогамией. В другом случае ядра амеб, напротив, гаплоидны, но в определенный период пара особей сливается, после чего образовавшаяся клетка с диплоидным ядром тут же делится путем мейоза. Интересно, что представители первой группы имеют лопастевидные, а второй – нитевидные ложноножки. Вероятно, эти амебы, несмотря на наличие сходных раковинок, не родственны между собой, и их объединение в один отряд является искусственным.
К отряду фораминифер (Foraminiferida) относят около 10 тысяч ныне живущих и еще примерно 20 тысяч ископаемых, известных по остаткам раковинок, видов корненожек. Фораминиферы отличаются тонкими ветвящимися ложноножками и имеют органическую, известковую, или сцементированную из песчинок раковинку. У примитивных форм она однокамерная, а у высших – многокамерная, разделенная на сообщающиеся порами отсеки. Форма раковинки у разных фораминифер может быть самой разнообразной – круглой, вытянутой, закрученной, напоминающей ягоду... Обычно ее размеры колеблются от 0,05 до 0,5 мм, однако в толще морских осадков встречаются трубчатые формы (например,Bathyosiphon) размером до нескольких сантиметров!