

Вопрос 94.
Вторичная структура ДНК.
Вторичная структура — конформационное расположение главной цепи макромолекулы (например, полипептидная цепь белка), независимо от конформации боковых цепей или отношения к другим сегментам . В описании вторичной структуры важным является определение водородных связей, которые стабилизируют отдельные фрагменты макромолекул. Это пространственная структура, образующаяся в результате взаимодействия между функциональными группами пептидного остова. Вторичная структура ДНК. Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. В 1953 г. Дж. Уотсон и Ф. Крик, обобщив работы многих современников (М. Уилкинс, Э. Чаргафф, А. Тода, Л. Полинг), описали вторичную структуру ДНК в виде двойной спирали. Она характерна для большинства молекул ДНК (в настоящее время известны и другие пространственные формы ДНК). Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8— 2,0 нм., две полинуклеотидные цепи антипараллельны друг другу т. е. направления образования фосфодиэфирных связей в них противоположны: в одной цепи 5’-З’, в другой — З’-5’. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары. Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого, а также между амидным и иминным атомами азота. Например, между аденином и тимином образуются две водородные связи и эти гетероциклические основания составляют комллементарную пару. Это означает, что адениновому основанию в одной цепи будет соответствовать тиминовое в другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин, между которыми возникают три водородные связи.
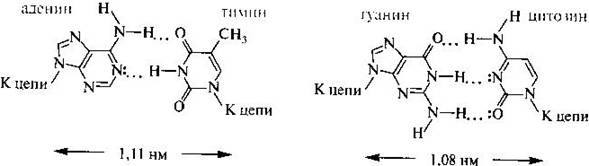
Связи между комплементарными основаниями — один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой. Это означает, что первичная структура, т. е. нуклеотидная последовательность одной цепи предопределяет первичную структуру второй цепи.
Пра́вила Ча́ргаффа — система эмпирически выявленных правил, описывающих количественные соотношения между различными типами азотистых оснований в ДНК. Были сформулированы в результате работы группы биохимика Эрвина Чаргаффа в 1949—1951 гг.
До работ группы Чаргаффа господствовала так называемая «тетрануклеотидная» теория, согласно которой ДНК состоит из повторяющихся блоков по четыре разных азотистых основания (аденин, тимин, гуанин и цитозин). Чаргаффу и сотрудникам удалось разделить нуклеотиды ДНК при помощи бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Они значительно отличались от эквимолярных, которых можно было бы ожидать, если бы все четыре основания были представлены в равных пропорциях. Соотношения, выявленные Чаргаффом для аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц), оказались следующими:
Количество аденина равно количеству тимина, а гуанина — цитозину: А=Т, Г=Ц.
Количество пуринов равно количеству пиримидинов: А+Г=Т+Ц.
Количество оснований с аминогруппами в положении 6 равно количеству оснований с кетогруппами в положении 6: А+Ц=Г+Т.
Вместе с тем, соотношение (A+Т):(Г+Ц) может быть различным у ДНК разных видов. У одних преобладают пары АТ, в других — ГЦ.
Вторичная структура РНК.
Молекулы РНК представляют собой единичные полинуклеотидные цепи. Отдельные участки молекулы РНК могут соединяться и образовывать двойные спирали. По своей структуре спирали РНК похожи на А-форму ДНК. Однако часто спаривание оснований в таких спиралях бывает неполным, а иногда даже и не уотсон-криковским. В результате внутримолекулярного спаривания оснований формируются такие вторичные структуры, как стебель-петля («шпилька») и псевдоузел. Вторичные структуры в мРНК служат для регуляции трансляции. Например, вставка в белки необычных аминокислот, селенометионина и пирролизина, зависит от «шпильки», расположенной в 3' нетранслируемой области. Псевдоузлы служат для программированного сдвига рамки считывания при трансляции.В вирусных мРНК сложные вторичные структуры (IRES) направляют трансляцию, независящую от узнавания кэпа и факторов инициации трансляции
Третичная структура ДНК. В клетках ДНК образует суперспирали, что обеспечивает компактность ее упаковки. ДНК длиной до 4 см располагается в хромосоме размером до 5 нм. Длина ДНК уменьшается в 100 тысяч раз. Третичная структура ДНК эукариот формируется путем взаимодействия с ядерными белками и на определенном этапе клеточного цикла приобретает форму хромосом.
Третичная структура Рнк (или трехмерная структура ]) — пространственное строение (включая конформацию) всей молекулы белка или другой макромолекулы, состоящей из единственной цепи.
Связь с первичной структурой.Третичная структура в значительной степени предопределена первичной структурой. Усилия по предсказанию третичной структуры белка основываясь на первичной структуре известна как задача предсказания структуры белка. Однако, окружающая среда, в которой белок сворачивается существенно определяет конечную форму, но обычно непосредственно не принимается во внимание текущими методами предсказания. Большинство таких методов полагаются на сравнения с уже известными структурами, и таким образом включают окружающую среду косвенно.
Стабилизирующие взаимодействия.В стабилизации третичной структуры белка принимают участие:
ковалентные связи (между двумя остатками цистеина — дисульфидные мостики);
ионные связи между противоположно заряженными боковыми группами аминокислотных остатков;
водородные связи;
гидрофильно-гидрофобные взаимодействия. При взаимодействии с окружающими молекулами воды белковая молекула «стремится» свернуться так, чтобы неполярные боковые группы аминокислот оказались изолированы от водного раствора; на поверхности молекулы оказываются полярные гидрофильные боковые группы.