

69. Образование о-гликозидов. Гликолиз гликозидов.
70.Окисление и восстановление моносахаридов. Восстановительные свойства альдоз. Восстановление моносахаридов.Восстановление. При восстановлении моносахаридов (их альдегидной или кетонной групп) образуются многоатомные спирты (полиолы) называемые альдитами. Эти кристаллические легко растворимые в воде вещества обладают сладким вкусом и часто используются как заменители сахара при сахарном диабете (ксилит, сорбит).

Восстановление моносахаридов проводят водородом в присутствии металлических катализаторов (палладий, никель). Шестиатомные спирты — глюцит (сорбит), дульцит и маннит — получаются при восстановлении соответственно глюкозы, галактозы и маннозы. Восстановление глюкозы в сорбит является одной из стадий промышленного синтеза аскорбиновой кислоты. При восстановлении альдоз получается лишь один полиол, кетоз — смесь двух полиолов, например, при восстановлении фруктозы борогидридом натрия NaВН4 образуются глюцит (сорбит) и маннит.

Окисление. Реакции окисления используют в структурных исследованиях и биохимических анализах для обнаружения моносахаридов, в частности глюкозы, в биологических жидкостях (моча, кровь). В зависимости от условий окисления образуются различные продукты. Окисление в щелочной среде. Альдозы в связи с наличием альдегидной группы способны восстанавливать в щелочкой среде катионы металлов (серебра, меди). Такие реакции возможны за счет таутомерного перехода в альдегидную форму, например реакция серебряного зеркала с реактивом Толленса. Кетозы тоже способны восстанавливать катионы металлов, так как они в щелочной среде изомеризуются в альдозы. Моносахариды в щелочной среде неустойчивы, поэтому при их
окислении получается смесь продуктов.

Моносахариды окисляются реактивом Бенедикта и реактивом Фелинга (фелингова жидкость), содержащими катионы меди(II). Различие между этими реактивами в том, что в фелинговой жидкости для стабилизации катионов меди(II) используют раствор калиево-натриевой соли винной кислоты (тартрат-ионы), а в реактиве Бенедикта — соли лимонной кислоты (цитрат-ноны). Принцип действия реактивов одинаков и основан на восстановлении двухвалентной меди в одновалентную с осаждением оксида меди(I) Сu2О (краснокирпичный цвет).

Реактивы Толленса, Бенедикта или Фелинга используются как качественные тесты для обнаружения альдоз и кетоз. Моносахариды и их производные (как и любые углеводы, вообще), вступающие в реакции с этими реактивами называют восстанавливающие. Гликозиды не проявляют восстановительной способности и не дают положительной пробы с этими реактивами. Окисление в нейтральной и кислой средах. Окисление альдоз без деструкции (разрушения) углеродного скелета проводят в нейтральной или кислой среде и получают различные кислоты. С помощью сильного окислителя — разбавленной азотной кислоты — концевые группы альдоз (альдегидная и первичноспиртовая) одновременно окисляются в карбоксильные группы, образуя альдаровые кислоты (называемые также сахарными). D-Глюкоза превращается при этом в глюкаровую кислоту.

Получающаяся из галактозы галактаровая (слизевая) кислота трудно растворима в воде, что используется для обнаружения галактозы методом окисления ее азотной кислотой. Мягкими окислителями (бромная вода) можно окислить альдегидную группу в карбоксильную не затрагивая других групп. При этом получаются альдоновые кислоты.

Альдоновые кислоты, являясь гидроксикислотами, выделяются в виде гамма- или дельта-лактонов, При окислении глюкозы бромной водой с высоким выходом получается глюконовая кислота, легко отщепляющая молекулу воды с образованием лактонов. В медицине используется кальциевая соль этой кислоты (глюконат кальция).

При окислении первичноспиртовой группы без затрагивания весьма склонной к окислению альдегидной группы получаются уроновые кислоты, синтез которых представляет собой сложную задачу. Обычно окислению подвергают моносахарид с защищенной альдегидной группой, например в виде гликозида. Уроновые кислоты, как и альдоновые, легко образуют лактоны. Уроновые кислоты при нагревании их солей с металлами (никель, магний) подвергаются декарбоксилированию. При этом из гексуроновых кислот образуются пентозы. В организме за счет ферментативной реакции декарбоксилирования глюкуроновой кислоты получается ксилоза.
Билет 71.
Природные
дисахариды: мальтоза,лактоза,сахароза,целлюлоза.Их
строение,цикла и оксо таутомерия и
биологическое значение.Восстанавливающие
и невосстанавливающие дисахариды.Дисахариды
состоят из двух моносахаридных остатков,
связанных гликозидной связью. Их можно
рассматривать как О-гликозиды, в которых
агликоном является остаток
моносахарида.Возможно два варианта
образования гликозидной связи:1) за счет
гликозидного гидроксила одного
моносахарида и спиртового гидроксила
другого моносахарида;2) за счет гликозидных
гидроксилов обоих моносахаридов.Дисахарид,
образованный первым способом, содержит
свободный гликозидный гидроксил,
сохраняет способность к цикло-оксо-таутомерии
и обладает восстанавливающими
свойствами.В
дисахариде, образованном вторым способом,
нет свободного гликозидного гидроксила.
Такой дисахарид не способен к
цикло-оксо-таутомерии и является
невосстанавливающим.В
природе в свободном виде встречается
незначительное число дисахаридов.
Важнейшими из них являются мальтоза,,
лактоза
и сахароза.Мальтоза
содержится
в солоде и образуется при неполном
гидролизе крахмала. Молекула мальтозы
состоит из двух остатков D-глюкозы в
пиранозной форме. Гликозидная связь
между ними образована за счет гликозидного
гидроксила в a-конфигурации одного
моносахарида и гидроксильной группы в
положении 4 другого моносахарида.

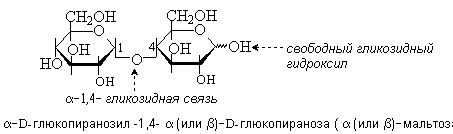
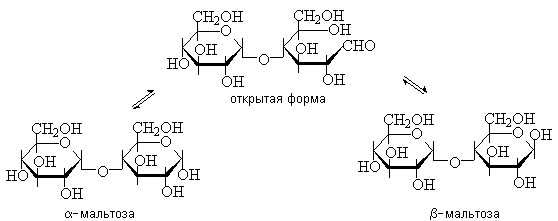
Мальтоза – это восстанавливающий дисахарид. Она способна к таутомерии и имеет a- и b-аномеры
Лактоза содержится в молоке (4-5%). Молекула лактозы состоит из остатков D-галактозы и D-глюкозы, связанных b-1,4-гликозидной связью. Лактоза – восстанавливающий дисахарид.

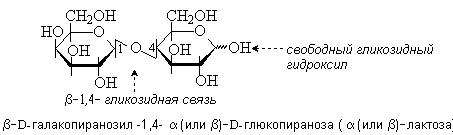
Сахароза содержится в сахарном тростнике, сахарной свекле, соках растений и плодах. Она состоит из остатков D-глюкозы и D-фруктозы, которые связаны за счет гликозидных гидроксилов. В составе сахарозы D-глюкоза находится в пиранозной, а D-фруктоза – в фуранозной форме. Сахароза – невосстанавливающий дисахарид
.

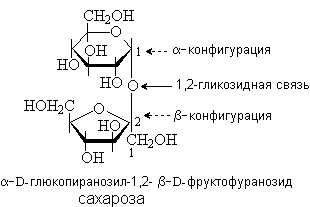
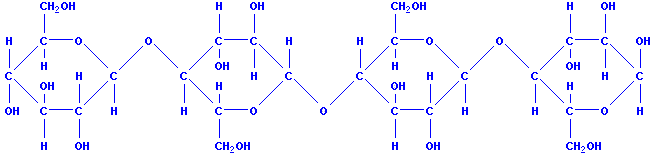
Целлюлоза тоже является природным полимером. Ее макромалекула состоит из многих остатков молекул глюкозы. Белое твердое вещество, нерастворимое в воде, молекула имеет линейное (полимерное) строение, структурная единица — остаток β-глюкозы [С6Н7О2(OH)3]n. Полисахарид, главная составная часть клеточных оболочек всех высших растений.
Роль сахарозы:Переваривание сахарозы начинается в тонком кишечнике.Кратковременное воздействие амилазы слюны существенной роли не играет,так как в просвете желудка кислая среда инактивирует этот фермент.В тонком кишечнике сахароза под действием фермента сахаразы,продуцируемой клетками кишечника,не выделяясь в просвет, а действуя на поверхности клеток (пристеночное пищеварение)Расщепление сахарозы приводит к высвобождению глюкозы и фруктозы.Проникновение моносахаридов через клеточные мембраны (всасывание)происходит путем облегченной диффузии при участии специальных транслоказ.Глюкоза всасывается еще и путем активного транспорта за счет градиента концентрации ионов натрия.Это обеспечивает ее всасывание даже при низкой концентрации в кишечнике.
МАЛЬТОЗЫ:
Мальтоза легко усваивается организмом человека. Расщепление мальтозы до двух остатков глюкозы происходит в результате действия фермента a-глюкозидазы, или мальтазы, которая содержится в пищеварительных соках животных и человека, в проросшем зерне, в плесневых грибах и дрожжах. Генетически обусловленное отсутствие этого фермента в слизистой оболочке кишечника человека приводит к врождённой непереносимости мальтозы — тяжёлому заболеванию, требующему исключения из рациона мальтозы, крахмала и гликогена или добавления к пище фермента мальтазы.
ЛАКТОЗЫ:
Биологическая роль лактозы такая же, как и у всех углеводов. В просвете тонкого кишечника под воздействием фермента лактазы она гидролизуется до глюкозы и галактозы, которые и усваиваются. Кроме того, лактоза облегчает всасывание кальция и является субстратом для развития полезных лактобактерий, которые составляют основу нормальной микрофлоры кишечника.
ЦЕЛЛЮЛОЗЫ:
целлюло́за
(клетчатка), полисахарид, полимер глюкозы. В клеточных стенках растений играют роль арматуры (каркаса), обеспечивая механическую прочность и эластичность растительных тканей. Особенно много целлюлозы в древесине – до 50%. Большинство животных не усваивает клетчатку, т.к. она, в отличие от крахмала, не расщепляется ферментом амилазой. Травоядные животные (жвачные) усваивают её благодаря целлюлазе – ферменту, вырабатываемому симбиотическими микроорганизмами, присутствующими в их желудке (гл. обр. в его отделе – рубце). Целлюлоза – один из самых распространённых природных полимеров.
Восстанавливающие дисахаридыВ этих дисахаридах один из моносахаридных остатков участвует в образовании гликозидной связи за счет гидроксильной группы (чаще всего при С-4). В дисахариде имеется свободная полуацетальная гидроксильная группа, вследствие чего сохраняется способность к раскрытию цикла.Восстановительные свойства таких дисахаридов и мутаротация их растворов обусловлены цикло-оксо-таутомерией.Представителями восстанавливающих дисахаридов являются мальтоза, целлобиоза, лактозаНевосстанавливающие:К этой группе принадлежит небольшое число дисахаридов, важнейшим из которых является сахароза.
В невосстанавливающих дисахаридах «вторая» молекула моносахарида в названии получает характерный для гликозидов суффикс –озид, например «фруктофуранозид» в сахарозе.
Наиболее характерной реакцией дисахаридов является кислотный гидролиз, приводящий к расщеплению гликозидной связи с образованием моносахаридов (во всех таутомерных формах). В общих чертах эта реакция аналогична гидролизу алкилгликозидов

Билет 72.
C3H5(OH)3+ Cu(OH)2 ----C3H5OHCuO2 +2 H2O (ярко-синий р-р глицерат меди):
( C6H10O5)n +J2 ----появляется синее окрашивание;
качест.р-ция на альдегиды: р-ция серебряного зеркала + Ag2O ------R-COOH +2Ag( блестящий налет);
Гомополисахариды (гликаны), состоящие из остатков одного моносахарида, могут быть гексозами или пентозами, то есть в качестве мономера может быть использована гексоза или пентоза. В зависимости от химической природы полисахарида различают глюканы (из остатков глюкозы), маннаны (из маннозы), галактаны (из галактозы) и другие подобные соединения. К группе гомополисахаридов относятся органические соединения растительного (крахмал, целлюлоза, пектиновые вещества), животного (гликоген, хитин) и бактериального (декстраны) происхождения.
Полисахари́ды — общее название класса сложных высокомолекулярных углеводов, молекулы которых состоят из десятков, сотен или тысяч мономеров — моносахаридов.Полисахариды необходимы для жизнедеятельности животных и растительных организмов. Они являются одним из основных источников энергии, образующейся в результате обмена веществ организма. Они принимают участие в иммунных процессах, обеспечивают сцепление клеток в тканях, являются основной массой органического вещества в биосфере.Классификация полисахаридовК полисахаридам относятся вещества, построенные из большого числа остатков моносахаридов или их производных. Если полисахарид содержит остатки моносахарида одного вида, его называют гомополисахаридом. В том случае, когда полисахарид составлен из моносахаридов двух видов или более, регулярно или нерегулярно чередующихся в молекуле, его относят к гетерополисахаридам.
К полисахаридам относятся, в частности:декстрин — полисахарид, продукт гидролиза крахмала;крахмал — основной полисахарид, откладываемый как энергетический запас у растительных организмов;гликоген — полисахарид, откладываемый как энергетический запас в клетках животных организмов, но встречается в малых количествах и в тканях растений;целлюлоза — основной структурный полисахарид клеточных стенок растений;хитин — основной структурный полисахарид экзоскелета насекомых и членистоногих, а также клеточных стенок грибов;галактоманнаны — запасные полисахариды некоторых растений семейства бобовых, такие как гуаран и камедь рожкового дерева;инулин - резервный углерод сложноцветных;глюкоманнан — полисахарид, получаемый из клубней конняку, состоит из чередующихся звеньев глюкозы и маннозы, растворимое пищевое волокно, уменьшающее аппетит;амилоид — применяется при производстве пергаментной бумаги;многоглюкоза — многоконечный продукт гидролиза большинства многосахаридов.
Билет 73.
Брожение углеводов, его применение. Молочнокислое брожение - процесс анаэробного окисления углеводов, конечным продуктом при котором выступает молочная кислота. Название получило по характеру продукта - молочной кислоте. Для молочнокислых бактерий является основным путём катаболизма углеводов и основным источником энергии в виде АТФ. Также молочнокислое брожение происходит в тканях животных в отсутствие кислорода при больших нагрузках. Спиртовое брожение.Спиртовое брожение — это процесс окисления углеводов, в результате которого образуются этиловый спирт, углекислота и выделяется энергия.
Сбраживание cахаров известно с глубокой древности. В течение столетий пивовары и виноделы использовали способность некоторых дрожжей вызывать спиртовое брожение, в результате которого сахара превращаются в спирт. Брожение производят главным образом дрожжи, а также некоторые бактерии и грибы. В различных странах для получения спирта используют различные микроорганизмы. Например, в Европе используют в основномдрожжи из рода Saccharomyces, в Южной Америке — бактерии Pseudomonas lindneri, в Азии — мукоровые грибы. Сбраживаться могут лишь углеводы, и притом весьма избирательно. Дрожжи сбраживают только некоторые 6-углеродные сахара (глюкозу, фруктозу, маннозу).
Билет 74. Классификация
По радикалу
Неполярные: глицин, аланин, валин, изолейцин, лейцин, пролин, метионин, фенилаланин, триптофан
Полярные незаряженные (заряды скомпенсированы) при pH=7: серин, треонин, цистеин, аспарагин, глутамин, тирозин
Полярные заряженные отрицательно при pH=7: аспартат, глутамат
Полярные заряженные положительно при pH=7: лизин, аргинин, гистидин
По функциональным группам
Алифатические
Моноаминомонокарбоновые: глицин, аланин, валин, изолейцин, лейцин
Оксимоноаминокарбоновые: серин, треонин
Моноаминодикарбоновые: аспартат, глутамат, за счёт второй карбоксильной группы несут в растворе отрицательный заряд
Амиды моноаминодикарбоновых: аспарагин, глутамин
Диаминомонокарбоновые: лизин, аргинин, несут в растворе положительный заряд
Серосодержащие: цистеин, метионин
Ароматические: фенилаланин, тирозин, триптофан, (гистидин)
Гетероциклические: триптофан, гистидин, пролин
Иминокислоты: пролин
По классам аминоацил-тРНК-синтетаз
Класс I: валин, изолейцин, лейцин, цистеин, метионин, глутамат, глутамин, аргинин, тирозин, триптофан
Класс II: глицин, аланин, пролин, серин, треонин, аспартат, аспарагин, гистидин, фенилаланин
Для аминокислоты лизин существуют аминоацил-тРНК-синтетазы обоих классов.

Билет 75.
Стериоизомерия аминокислот. Кислотно-основные свойства , биополярная структура аминокислот. Аминокислотами называются органические карбоновые кислоты, у которых как минимум один из атомов водорода углеводородной цепи замещен на аминогруппу. В зависимости от положения группы —NН2 различают α, β, γ и т. д. L-аминокислоты. К настоящему времени в различных объектах живого мира найдено до 200 различных аминокислот. В организме человека содержится около 60 различных аминокислот и их производных, но не все они входят в состав белков. Кислотно-основные свойства аминокислот связаны с наличием в их структуре двух ионизируемых групп-карбоксильной и аминогруппы, поэтому амнокислоты могут проявлять свойства как кислот, так и оснований, т.е. они являются амфотерными соединениями. В кристаллическом состоянии и в водных растворах a-аминокислоты существуют в виде биполярных ионов, называемых также цвиттерионами. Ионное строение обуславливает некоторые особенности свойств a-аминокислот: высокую температуру плавления (200-300°С), нелетучесть, растворимость в воде и нерастворимость в неполярных органических растворителях. С растворимостью аминокислот в воде связана их всасываемость и транспорт в организме. Ионизация молекул аминокислот зависит от рН раствора.
Билет 77.
Качественные реакции на аминокислоты.
Особенность химии аминокислот и белков заключается в использовании многочисленных качественных реакций.
Общая качественная реакция альфааминокислот – реакция с нингидрином.Продукт нингидридной реакции имеет сине-фиолетовый цвет, что используется для визуального обнаружения аминокислот на хроматограммах, а также спектрофотометрического определения с помощью аминокислотных анализаторов(продукт поглощает свет в области 550- 570 нм).
Для обнаружения пептидных связей в пептидах и белках служит биуретовая реакция,в которую вступают все пептиды и белки , содержащие по крайней мере две такие связи.
Существует также ряд частных реакций, позволяющих обнаруживать отдельные альфааминокислоты или группы родственных альфааминокислот.
Триптофан обнаруживают при помощи реакции с n-диметиламинобензаль дегидом в среде серной кислоты по появляющемуся красно-фиолетовому окрашиванию. Эта реакция используется для количественного анализа триптофана в продуктах расщепления белков.
Цистеин обнаруживается с помощью нескольких качественных реакций, основанных на реакционной способности содержащейся в нем меркаптогруппы. Например , при нагревании раствора белка с ацетатом свинца в щелочной среде образуется черный осадок сульфида свинца, что указывает на присутствие в белках цистеина.
Для обнаружения ароматических и гетероциклических альфааминокислот используется ксантопротеиновая реакция.Например, при действии концентрированной азотной кислотой на тирозин образуется нитросоединение , окрашенное в желтый цвет. При добавлении к нему щелочи окраска становится оранжевой в связи и ионизацией фенольной гидроксильной группы и увеличением вклада аниона в сопряжение.( сама печатала, возможно наличие ошибок, ребят)
Билет 79
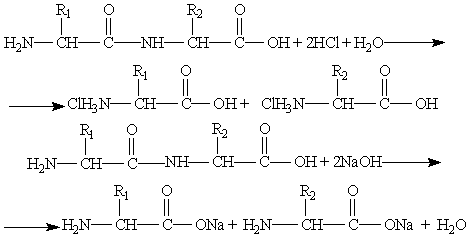
Способность к гидролизу – это основное свойство пептидов. При гидролизе происходит частичное или полное разделение пептидной цепи и получаются более короткие пептиды или α-аминокислоты, которые составляют цепь. При анализе продуктов полного гидролиза, можно установить из каких аминокислот состоит пептид. Полный гидролиз получается, если долго нагревать пептид с концентрированной соляной кислотой. Путем частичного гидролиза пептида можно установить последовательность аминокислот в цепи. Гидролиз пептидов можно проводить в щелочной и кислой среде, а также под действием ферментов. Соли аминокислот получаются в щелочной и кислой средах. Важность ферментативного гидролиз заключается в том, что он позволяет разделять строго определенные участки пептидной цепи. Многие пептиды могут проявляют биологическую активность. В кислой и щелочной средах образуются соли аминокислот:
Изоэлектрической точкой называют такую концентрацию водородных ионов, т. е. такое значение рН, при котором имеется равновесная концентрация положительных и отрицательных зарядов у белковой молекулы. При изоэлектрической точке молекула белка электронейтральна и не передвигается в электрическом поле. В таком состоянии молекулы белка легко выпадают в осадок. Большинство белков имеют изоэлектрическую точку при рН от 4,0 до 8,0ю Большенство белков в клетке имеет в своем составе больше анионогенных групп (-СОО-), то изоэлектрическая точка этих белков лежит в слабокислой среде. Изоэлектрическая точка белков, в составе которых преобладает катиогогенные группы , находится в щелочной среде.(Пример аригин и лизин –гистоны ,входящие в состав хроматина).
Качественная реакция на белок – биуретовая реакция, которая обусловлена наличием пептидных связей в белковых молекулах. В щелочной среде белки образуют с ионами меди растворимое комплексное соединение сине-фиолетового цвета.
Билет 80.
Простые: сюда входят сложные эфиры жирных кислот и спирты. 1. Глицериды – это сложные эфиры 3-атомного спирта глицерина и высших жирных кислот. 2. Воски – сложные эфиры 1- или 2-атомных спиртов и высших жирных кислот.Сложные: сложные эфиры жирных кислот со спиртами, в которые включены и иные группы. 1. Фосфолипиды. В этих жирах кроме жирных кислот и спирта включены и следы фосфорной кислоты, азотистые компоненты, а также сфинголипиды и глицерофосфолипиды. 2. Гликолипиды 3. Стероиды 4. Иные сложные жиры: аминолипиды, сульфолипиды, а также липопротеины.
Омыляемые липиды. Структурные компоненты омыляемых липидов связаны сложноэфирной связью. Эти липиды легко гидролизуются в воде под действием щелочей или ферментов. Омыляемые липиды включают три группы веществ: сложные эфиры, фосфолипиды и гликолипиды. В группу сложных эфиров входят нейтральные жиры (глицерин+три жирные кислоты), воски (жирный спирт+жирная кислота) и эфиры стеринов (стерин+жирная кислота). Группа фосфолипидов включает фосфатидовые кислоты (глицерин+две жирные кислоты+фосфатная группа), фосфатиды (глицерин+две жирные кислоты+фосфатная группа+спирт) и сфинголипиды (сфингозин+жирная кислота+фосфатная группа+спирт). К группе гликолипидов относятся цереброзиды (сфингозин+жирная кислота+один углеводный остаток) и ганглиозиды (сфингозин+жирная кислота+несколько углеводных остатков, в том числе нейраминовая кислота). Группа неомыляемых липидов включает предельные углеводороды и каротиноиды, а также спирты. В первую очередь это спирты с длинной алифатической цепью, циклические стерины (например, холестерин) и стероиды (эстрадиол, тестостерон и др.). Важнейшую группу липидов образуют жирные кислоты. К этой группе относятся также эйкозаноиды, которые можно рассматривать как производные жирных кислот.
Пальмитиновая кислота- CH3- CH2(14 раз)-СООН
Пальмитолеиновая кислота (С15Н29СOOH) или (СН3-(СН2)5-СН=СН-(СН2)7-СООН)
Стеариновая кислота(С17Н35СООН) или CH3-CH2(16раз)-COOH
Линолевая кислота (CH3(CH2)3-(CH2CH=CH)2(CH2)7COOH ) или С17H31COOH
Линоленовая кислота (CH3(CH2CH=CH)3(CH2)7COOH)
Олеиновая кислота СН3(СН2)7СН=СН(СН2)7СООН или C17H33COOH
Арахидоновая кислота CH3(CH2)4(CH=CHCH2)4(CH2)2COOH или C20H32O2
(Обозначение числа и конформация ВЖК не нашел)
Билет 81.
Это
сложные эфиры жирных кислот и спиртов.
Они образуются за счет реакции
этерификации.
![]()
Все простые липиды делятся на три группы: воска ,жиры, церамид
(Физические свойства ПРОСТЫХ ЛИПИДОВ не нашел)
Во́ски — распространённые в растительном и животном мире сложные эфиры высших жирных кислот и высших высокомолекулярных спиртов. Очень устойчивы, нерастворимы в воде, но хорошо растворимы в бензине, хлороформе, эфире. По происхождению воски можно разделить на животные: пчелиный вырабатывается пчёлами; шерстяной (ланолин) предохраняет шерсть и кожу животных от влаги, засорения и высыхания; спермацет добывается из спермацетового масла кашалотов; растительные воски покрывают тонким слоем листья, стебли, плоды и защищают их от размачивания водой, высыхания, вредных микроорганизмов, иногда в качестве резервных липидов входят в состав семян , ископаемый воск (озокерит) состоит главным образом из предельных углеводородов.
1.Свечи
2.Изготовление выплавляемых моделей для литья, так называемых восковок.
3. В натуральной косметике — загуститель для кремов, мазей и лубрикантов, основной компонент помад, твердых духов
4.В составе вара.
5.Компонент политур и мастик для полировки и защиты мебели, деревянных изделий, паркетных полов, мрамора и др.
6.Гемостатический костный воск используется для остановки кровотечения из кости и широко применяется в нейрохирургии. Впервые разработан в начале ХХ века британским хирургом Виктором Горслеем
7. Один из компонентов, необходимый для воскографии (в изобразительном искусстве).
8.Для защиты продуктов питания (фруктов, сыров) при транспортировке.
Билет 82.
Жиры, или триглицериды — природные органические соединения, полные сложные эфиры глицерина и одноосновных жирных кислот; входят в класс липидов. В живых организмах выполняют структурную, энергетическую и др. функции. Наряду с углеводами и белками, жиры — один из главных компонентов питания. Состав жиров отвечает общей формуле: CH2-O-C(O)-R¹ | CH-О-C(O)-R² | CH2-O-C(O)-R³, где R¹, R² и R³ — радикалы (иногда различных) жирных кислот.
Насыщенные:
лауриновая (C11H23COOH)
миристиновая (C13H27COOH)
стеариновая (C17H35COOH)
пальмитиновая (C15H31COOH)
Ненасыщенные:
пальмитолеиновая (C15H29COOH, 1 двойная связь)
олеиновая (C17H33COOH, 1 двойная связь)
линолевая (C17H31COOH, 2 двойные связи)
линоленовая (C17H29COOH, 3 двойные связи)
арахидоновая (C19H31COOH, 4 двойные связи, реже встречается)
Чаще всего в животных жирах встречаются стеариновая и пальмитиновая кислоты, ненасыщенные жирные кислоты представлены в основном олеиновой, линолевой и линоленовой кислотами. Физико-химические и химические свойства жиров в значительной мере определяются соотношением входящих в их состав насыщенных и ненасыщенных жирных кислот.
В растениях жиры содержатся в сравнительно небольших количествах, за исключением семян масличных растений, в которых содержание жиров может быть более 50 %. Насыщенные жиры расщепляются в организме на 25—30 %, а ненасыщенные жиры расщепляются полностью.
Йодное число является простой и быстро определяемой химической постоянной для жиров или масел. Это важный показатель жиров, характеризующий содержание ненасыщенных соединений, но не определяющий отдельные жирные кислоты. Йодное число выражается количеством граммов йода, которое может присоединиться по двойным связям к 100 г жира или другого исследуемого продукта. Для определения йодного числа применяют растворы хлорида йода - ICl, бромида йода - IBr или йода в растворе сулемы, которые обладают большей реакционной способностью, чем растворы йода в спирте или других растворителях. Йодное число является мерой ненасыщенности кислот жиров. Оно особенно важно для оценки качества высыхающих масел.
Билет 85.
неомыляемые липиды. К ним относятся стероидные гормоны, простаноиды, некоторые коферменты, жирорастворимые витамины, объединяемые одним названием — низкомолекулярные биорегуляторы.Под этим названием объединяют ряд углеводородов и их кислородсодержащих производных, в состав которых входят гидроксильная, карбонильная группы, углеродный скелет которых построен из двух, трех и более звеньев изопрена. Сами углеводороды называют терпеновыми углеводородами, а их кислородсодержащие производные — терпеноидами. Терпенами богаты эфирные масла растений (герани, розы, лаванды, лимона, перечной мяты и др.), смола хвойных деревьев и каучуконосов. К терпенам относятся и различные растительные пигменты и жирорастворимые витамины. Группировка терпенового типа (изопреноидная цепь) включена в структуру многих биологически активных соединений. В большинстве терпенов изопреновые фрагменты соединены друг с другом по принципу «голова к хвосте — изопреновое правило (Л. Ружичка, 1921), как показано на примере мирцена.

Терпеновые углеводороды и терпеноиды. Общая формула большинства терпеновых углеводородов — (С5Н8)n. Они могут иметь ациклическое и бициклическое (би-, три- и полициклическое строение). С учетом числа изопреновых группировок в молекуле различают • монотерпены (две изопреновые группировки); • сесквитерпены (три изопреновые группировки); • дитерпены (четыре изопреновые группировки); • тритерпены (шесть изопреновых группировок); • тетратерпены (восемь изопреновых группировок). Примерами ациклических терпенов служат упоминавшийся выше мирцен — монотерпен, содержащийся в эфирных масла хмеля и благородного лавра; родственный мирцену спирт гераниол входящий в состав эфирных масел герани и розы и при мягком окислении образующий альдегид — цитраль а.

Лимонен — представитель моноциклических терпенов. Он содержится в лимонном масле и скипидаре. Лимонен входит в состав масла тмина. Рацемическая форма лимонена (дипентен) может быть получена в результате реакции диенового синтеза из изопрена при нагревании.

При восстановлении оптически активного лимонена или дипентена получается ментан, а при полной их гидратации в кислой среде, протекающей в соответствии с правилом Марковникова образуется двухатомный спирт терпин. Терпин в виде гидрата применяется как отхаркивающее средство при хроническом бронхите.

3амещенные дипентены (например, каннабидиол) — психоактивное начало гашиша (марихуаны). Ментол — имеет скелет ментана подобно лимонену. Ментол содержится в эфирном масле перечной мяты. Из приведенной конформации ментола видно, что все три заместителя в циклогексановом кольце занимают экваториальные положения.

для получения рацемического ментола в промышленности м-крезола алкилируют изопропиловым спиртом в присутствии серной кислоты. При этом из изопропилового спирта первоначально генерируется изопропильный катион — электрофильная частица. При последующем восстановлении продукта алкилирования — тимола получается ментол.

Ментол оказывает антисептическое, успокаивающее и болеутоляющее (отвлекающее) действие, входит в состав валидола, а также мазей, применяемых при насморке. Альфа-пинен — бициклический монотерпен ряда пинана. Его левовращающий энантиомер — важная составная часть скипидара, получаемого из хвойных деревьев.
Камфора — бициклический кетон — редкий пример соединения, в котором шестичленный цикл имеет конформацию ванны. Камфора содержит два асимметрических атома углерода, и для нее следовало бы ожидать существования четырех оптически активных стереоизомеров. Их, однако, известно всего два, так как вследствие жесткости системы конфигурация обоих хиральных центров может измениться лишь одновременно (транс-расположение мостиковой группы -С(СН3)2- между узловыми асимметрическими атомами невозможно).

Камфора издавна применяется в медицине как стимулятор сердечной деятельности. Ее правовращающий стереоизомер выделяют из эфирного масла камфорного дерева. При действии брома на камфору замещается альфа-положение по отношению к карбонильной группе с образованием бромокамфоры.

Бромокамфора улучшает деятельность сердца, оказывает успокаивающее действие на центральную нервную систему.
Особую группу терпенов составляют каротиноиды — растительные пигменты. Некоторые из них играют роль витаминов или предшественников витаминов, а также участвуют в фотосинтезе. Большинство каротиноидов относится к тетратерпенам. Их молекулы содержат значительное число сопряженных двойных связей, поэтому имеют желто-красную окраску. Для природных каротиноидов характерна транс-конфигурация двойных связей. Каротин — растительный пигмент цвета, в большом количестве содержавшийся в моркови, а также томатах и сливочном масле. Известны три его изомера, называемые альфа-, бета- и гамма-каротинами, различающиеся числом циклов и положением двойных связей. Все они являются предшественниками витаминов группы А. Молекула бета-каротина симметрична и состоит из двух одинаковых частей.

СТЕРИНЫ, вещества, относящиеся к группе липоидов, входящие в состав неомыляемой части жиров и имеющие большое распространение в животном (зоостерины) и растительном (фито-стерины) мире. Различные С. сопутствуют друг другу, что затрудняет выделение отдельных С, в чистом виде.
Стеринами называются cтероидные спирты. Все стерины содержат β-гидроксильную группу при С-3 и одну или несколько двойных связей в кольце В и боковой цепи. В молекулах стеринов отсутствуют карбоксильные и карбонильные группы.
В организме животных наиболее важным стерином является холестерин. В растениях и микроорганизмах содержится множество родственных соединений, например эргостерин, β-ситостерин, стигмастерин.
Холестерин присутствует во всех животных тканях, особенно в нервных тканях. Он является важнейшей составной частью клеточных мембран, где регулирует их текучесть (см. с. 219). Запасной и транспортной формами холестерина служат его эфиры с жирными кислотами. Наряду с другими липидами холестерин и его эфиры присутствуют в составе липопротеидных комплексов плазмы крови (см. с. 273). Холестерин входит в состав желчи и многих желчных камней.
Стеринами называются cтероидные спирты. Все стерины содержат β-гидроксильную группу при С-3 и одну или несколько двойных связей в кольце В и боковой цепи. В молекулах стеринов отсутствуют карбоксильные и карбонильные группы.
В организме животных наиболее важным стерином является холестерин. В растениях и микроорганизмах содержится множество родственных соединений, например эргостерин, β-ситостерин, стигмастерин.
Холестерин присутствует во всех животных тканях, особенно в нервных тканях. Он является важнейшей составной частью клеточных мембран, где регулирует их текучесть (см. с. 219). Запасной и транспортной формами холестерина служат его эфиры с жирными кислотами. Наряду с другими липидами холестерин и его эфиры присутствуют в составе липопротеидных комплексов плазмы крови (см. с. 273). Холестерин входит в состав желчи и многих желчных камней.
Сюда перенаправляется запрос «Витамин А». На эту тему нужна отдельная статья.
![]()
Структурная формула молекулы ретинола
Ретино́л (истинный витамин A, транс-9,13-Диметил-7-(1,1,5-триметилциклогексен-5-ил-6)-нонатетраен-7,9,11,13-ол) — жирорастворимый витамин, антиоксидант. В чистом виде нестабилен, встречается как в растительных продуктах, так и в животных источниках. Поэтому производится и используется в виде ретинола ацетата и ретинола пальмитата. В организме синтезируется из бета-каротина. Необходим для зрения и костей, здоровья кожи и волос, нормальной работы иммунной системыи т.д.
Витамин E — группа жирорастворимых биологически активных веществ (токоферолы итокотриенолы), проявляющих антиоксидантные свойства.
Витамин К относится к группе липофильных (жирорастворимых) и гидрофобных витаминов, необходимых для синтеза белков, обеспечивающих достаточный уровень коагуляции. Химически, является производным 2-метил-1,4-нафтохинона. Играет значительную роль в обмене веществ в костях и в соединительной ткани, а также в здоровой работе почек. Во всех этих случаях витамин участвует в усвоении кальция и в обеспечении взаимодействия кальция и витамина D. В других тканях, например, в лёгких и в сердце, тоже были обнаружены белковые структуры, которые могут быть синтезированы только с участием витамина К.
стероидные гормоны- (половые, глюко- и минералокортикоиды),
Строение холестерина
Холестерин — это натуральное вещество, вырабатываемое нашим организмом, а также содержащееся в пище, которую мы едим. Оно необходимо для жизни — и вместе с тем излишек холестерина может быть смертелен. Немногие элементы нашей пищи удостаивались в последнее время такого повышенного внимания, как холестерин. Многие люди осознают, насколько вредно для их организма повышенное содержание холестерина в крови, непосредственно связанное с сердечными заболеваниями, представляющими наибольшую угрозу для жизни. Тем не менее они не знают, какие продукты (и особенно какие жиры) повышают уровень холестерина и почему овсяные отруби полезны, а сливки, масло, жареные продукты, заправки к салатам или авокадо — наоборот. Другое всеобщее незнание касается разницы между "полезным" и "вредным" холестерином и способов регулирования его содержания в крови с помощью диеты

Эйкозаноиды — окисленные производные полиненасыщенных жирных кислот — эйкозотриеновой (С20:3), арахидоновой (эйкозотетраеновая, С20:4), тимнодоновой (эйкозопентаеновая, С20:5). Пищевыми источниками полиненасыщенных жирных кислот являются растительные масла, рыбий жир и препараты омега-3-жирных кислот.
Депонироваться эйкозаноиды не могут, разрушаются в течение нескольких секунд, поэтому клетка должна синтезировать их постоянно из поступающих в неё соответствующих жирных кислот.
Выделяют три основные группы эйкозаноидов:
простагландины
лейкотриены
тромбоксаны
Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов. Выделяют типы простагландинов A, B, C, D, E, F. Их функции сводятся к изменению тонуса гладких мышц бронхов, мочеполовой и сосудистой систем, желудочно-кишечного тракта, при этом направленность изменений различна в зависимости от типа простагландинов и условий. Они также влияют на температуру тела.
Билет 86.
Нуклеиновые кислоты (ДНК и РНК) относятся к сложным высокомолекулярным соединениям, состоят из небольшого числа индивидуальных химических компонентов более простого строения. Так, при полном гидролизе нуклеиновых кислот (нагревание в присутствии хлорной кислоты) в гидролизате обнаруживают пуриновые и пиримидиновые основания, углеводы (рибоза и дезоксирибоза) и фосфорную кислоту
В молекуле ДНК углевод представлен дезоксирибозой, а в молекуле РНК – рибозой, отсюда их названия: дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК) кислоты. Кроме того, они содержат фосфорную кислоту, по два пуриновых и по два пиримидиновых основания; различия только в пиримидиновых основаниях: в ДНК содержится тимин, а в РНК – урацил. В составе ДНК и РНК открыты так называемые минорные (экзотические) азотистые основания (строение некоторых из них приводится далее).



Билет 87.
Азо́тистые основа́ния — гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеиновых кислот. Для сокращенного обозначения пользуются большими латинскими буквами. К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C), которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил (U) встречается только в РНК.[1]
Аденин и гуанин являются производными пурина, а цитозин, урацил и тимин — производными пиримидина.
Тимин, который присутствует только в ДНК, и урацил, который встречается только в РНК, обладают сходной химической структурой. Урацил отличается от тимина отсутствием метильной группы у 5-го атома углерода.
Азотистые основания, соединяясь ковалентной связью с 1' атомом рибозы или дезоксирибозы, образуют N-гликозиды, которые называют нуклеозиды. Нуклеозиды, в которых к 5'-гидроксильной группе сахара присоединены одна или несколько фосфатных групп, называются нуклеотидами. Эти соединения являются строительными блоками молекул нуклеиновых кислот — ДНК и РНК.
После образования молекулы нуклеиновой кислоты входящие в её состав азотистые основания могут вступать в различные химические реакции под действием ферментов а также факторов внешней среды. Таким образом, нуклеиновые кислоты часто содержат модифицированные азотистые основания. Типичной модификацией такого рода является метилирование
Билет 88.
Аденин
В результате дезаминирования аденина образуется гипоксантин, который имеет структуру, сходную с таутомером аденина, и спаривается с цитозином вместо тимина. Происходит транзиция A-T в G-C.
ДЕЗАМИНИРОВАНИЕ, отщепление аминогрупп (в форме NH3 или свободного N).—Д. аминокислот и белков хим. путем достигается действием азотистой кислоты. При этом алифатические NHj-группы аминокислот реагируют по схеме: R, NHa+HN02->R. ОН + N2 +ЩО.
Дезаминирование гуанина(превращение его в ксантин)
Дезаминирование аденина в составе поли-нуклеотида (превращение в гипоксантин) меняет информац. смысл и приводит к точковой мутации. Дезаминирование гуанина (превращение его в ксантин) в составе матричных полинуклеотидов приводит к блокированию репликации и транскрипции. Метилирование П. о. по N-7 в составе матричных полинуклеотидов не сопровождается изменением генетич. смысла основания.
П. о. представляют собой высокоплавкие (т. пл. > 250 °С), бесцв. кристаллич. соед., плохо раств. в горячей воде (особенно гуанин), не раств. в этаноле и диэтиловом эфире. Содержание редких таутомерных форм (иминотаутоме-ры А и G по С-6 и С-2 соотв., енольного таутомера G по С-6) не превышает в норм. условиях 10-3%. Протонирование и депрото-нирование П. о. сопровождается изменениями УФ спектров поглощения (см. табл.) и реакц. способности.

.дезаминирование аденина, цитозина и гуанина...
Билет
89.Нуклеиновые
кислоты –
это линейные полимеры нуклеотидов,
состоящих из пуриновых и пиримидиновых
оснований (нуклеиновых оснований),
углевода и фосфата.Азотистые
основания:
пуриновые
основания - аденин (А), гуанин
(Г);
пиримидиновые основания - цитозин
(Ц), урацил (У), тимин (Т)В состав
мононуклеотидов обычно входят углеводы:
D-рибоза, глюкоза (редко) и
D-дезоксирибоза.
Нуклеиновые кислоты
разделены на два класса: рибонуклеиновые
(РНК) и дезоксирибонуклеиновые (ДНК), в
зависимости от углевода.Описание
последовательности нуклеотидов в
нуклеиновой кислоте отражает ее первичную
структуру. Расположение длинной
полинуклеотидной цепи в пространстве
отражается в ее вторичной структуре.В
молекулах ДНК (согласно модели Дж.Уотсона
и Ф.Крика) полинуклеотидная цепь
спирализована в правую спираль с периодом
идентичности 3,4 нм и расстоянием между
плоскостями оснований 0,34 нм. Две цепи
сплетены друг с другом в закрученную
вокруг одной оси двойную спираль так,
что на каждый виток спирали приходится
10 пар оснований; диаметр спирали 2,0 нм.
Обе цепи удерживаются друг около друга
за счет водородных связей между
основаниями, находящимися внутри двойной
спирали.Особенностью ДНК является
эквивалентность количеств пуриновых
оснований количествам пиримидиновых
оснований, то есть отношения аденин –
тимин, гуанин – цитозин всегда равны
единице. Это соотношение объясняется
тем, что пространственная (вторичная)
структура ДНК создается за счет
образования водородных связей между
остатками нуклеиновых оснований таким
образом, что аденин (А) связывается
только с тимином (Т), а гуанин (Г) – только
с цитозином (Ц).Парные
основания получили
название комплементарных: А-Т; Г-Ц.
Комплементарность нуклеиновых оснований
лежит в основе корреляции между структурой
и функцией нуклеиновых
кислот.КОМПЛЕМЕНТАРНОСТЬ в химии,
пространственное
соответствие структур двух молекул (разных
или одинаковых), благодаря к-рому возможно
образование между ними водородных
связей и
осуществление межмол. взаимодействий.
В широком смысле также взаимное
соответствие противоположных
электростатич. зарядов на молекулах и
энергий сопряженных р-ций. В последнем
случае рассматриваются параллельно
протекающие р-ции, связанные друг с
другом так, что стадия, сопровождающаяся
выделением энергии, сопряжена со стадией,
для осуществления к-рой необходимо
потребление энергии. Наиб. распространено,
особенно в биохимии и
биоорг. химии,
понятие структурной комплементарности.
Благодаря этому виду комплементарности,
к-рое осуществляется по принципу "ключ
- замок", образуются
комплексы антиген - антитело, фермент - субстрат, четвертичная
структура белков,
вторичная и третичная структура
нуклеиновых к-т. В последнем случае
комплементарность проявляется особенно
ярко. Комплементарность аденина тимину игуанина цитозину (в
узком смысле термин "комплементарность"
иногда употребляется именно для этого
случая) открыта Дж. Уотсоном и Ф. Криком
в 1953 и легла в основу построенной ими
модели двойной спирали ДНК. Данный
тип комплементарности реализуется
благодаря образованию водородных
связей между
протонодонорными и протоноакцепторными
группами в азотистых
основаниях (см.
рис.). При
 Водородные
связи (обозначены
точками) между комплементарными основаниями,
входящими в ДНК и РНК;
R остаток фосфорилированной пентозы. этом
образуются специфич. пары комплементарных оснований,
имеющие почти одинаковые размеры.
Поэтому двойная спираль имеет очень
однородную регулярную структуру, мало
зависящую от конкретной
последовательностиоснований -
св-во очень важное для обеспечения
универсальности
механизмов репликации (самовоспроизведение ДНК или РНК),транскрипции (синтез РНК на
ДНК-матрице) и трансляции (синтез
белков на
РНК-матрице). В каждом из этих т.н.
матричных процессов комплементарность
играет определяющую роль. Напр.,
при трансляции важное
значение имеет комплементарность между
тройкойоснований матричной
РНК (т.н. кодоном,
см. Генетический
код)
и тройкой оснований транспортной
РНК (поставляют
во времятрансляции аминокислоты).
Комплементарность определяет также
вторичную структуру нуклеиновых к-т.
Одноцепочечные РНКблагодаря
комплементарности оснований,
навиваясь сами на себя, образуют
относительно короткие двухспиральные
области ("шпильки"
и "петли"), соединенные одноцепочечными
участками. Комплементарность в
отдельных парах оснований ДНК может
нарушаться из-за появления отклонений
в их строении, к-рые могут возникать
спонтанно или в результате действия
разл. факторов (химических и физических).
Следствием этих изменений м.
б. мутации. Комплементарность
лежит в основе мн. явлений биол.специфичности,
связанных с "узнаванием" на мол.
уровне, -ферментативного катализа,
самосборки биол.
структур, высокой точности передачи
генетич. информации и др.
Водородные
связи (обозначены
точками) между комплементарными основаниями,
входящими в ДНК и РНК;
R остаток фосфорилированной пентозы. этом
образуются специфич. пары комплементарных оснований,
имеющие почти одинаковые размеры.
Поэтому двойная спираль имеет очень
однородную регулярную структуру, мало
зависящую от конкретной
последовательностиоснований -
св-во очень важное для обеспечения
универсальности
механизмов репликации (самовоспроизведение ДНК или РНК),транскрипции (синтез РНК на
ДНК-матрице) и трансляции (синтез
белков на
РНК-матрице). В каждом из этих т.н.
матричных процессов комплементарность
играет определяющую роль. Напр.,
при трансляции важное
значение имеет комплементарность между
тройкойоснований матричной
РНК (т.н. кодоном,
см. Генетический
код)
и тройкой оснований транспортной
РНК (поставляют
во времятрансляции аминокислоты).
Комплементарность определяет также
вторичную структуру нуклеиновых к-т.
Одноцепочечные РНКблагодаря
комплементарности оснований,
навиваясь сами на себя, образуют
относительно короткие двухспиральные
области ("шпильки"
и "петли"), соединенные одноцепочечными
участками. Комплементарность в
отдельных парах оснований ДНК может
нарушаться из-за появления отклонений
в их строении, к-рые могут возникать
спонтанно или в результате действия
разл. факторов (химических и физических).
Следствием этих изменений м.
б. мутации. Комплементарность
лежит в основе мн. явлений биол.специфичности,
связанных с "узнаванием" на мол.
уровне, -ферментативного катализа,
самосборки биол.
структур, высокой точности передачи
генетич. информации и др.
Билет 90.
Присоединение к нуклеозиду одного остатка фосфорной кислоты приводит к образованию нуклеозидмонофосфата (НМФ), или нуклеотида. Фосфорная кислота взаимодействует с пятым атомом углерода рибозы или дезоксирибозы. Таким образом, нуклеотиды являются фосфорными эфирами нуклеозидов.Из нуклеозидов, углеводным компонентом которых является рибоза, образуются рибонуклеотиды:Аденозинмонофосфат (АМФ, или адениловая кислота); гуанозинмонофосфат (ГМФ, или гуаниловая кислота); цитидинмонофосфат (ЦМФ, или цитидиловая кислота); уридинмонофосфат (УМФ, или уридиловая кислота). Нуклеознды, построенные на основе дезоксирибозы, в результате присоединения остатка фосфорной кислоты образуют дезоксирибонуклеотиды:Дезоксиаденозинмонофосфат (дАМФ, или дезоксиадениловая кислота);Дезоксигуанозинмонофосфат (дГМФ, или дезоксигуаниловая кислота);Дезоксицитидинмонофосфат (дЦМФ, или дезоксицитидиловая кислота);Дезокситимидинмонофосфат (дТМФ, или дезокситимидиловая кислота).Строение нуклеотидов можно представить на примере структурных формул аденозинмонофосфата и дезоксици-динмонофосфата.Нуклеотиды выполняют ряд важнейших биологических функций. Прежде всего, они образуют нуклеиновые кислоты. Кроме этого, нуклеотиды входят в состав некоторых ферментов в качестве коферментов, а также участвуют в построении производных соединений, в которых аккумулируется энергия, Необходимая для протекания процессов жизнедеятельности в клетке.

Строение нуклеозидов(изображение)

Отличия основания, нуклеозида, нуклеотида.
Нуклеоти́ды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
Билет 91.
Циклические пуриновые нуклеотиды — аденозин-3´, 5´-монофосфат (цАМФ) и гуанозин-3´, 5´-монофосфат (цГМФ) — играют важную физиологическую роль практически во всех видах обмена веществ в клетке в их фенотипическом проявлении. Помимо этого, имеются прямые указания на участки аденилциклазной системы в контроле механизмов генетической информации за счет изменения активности ферментов, регулирующих фосфорилирование гистонов. Обнаружено также участие цАМФ в формировании иммунного ответа и механизмах фагоцитоза. Следовательно, многообразие физиологических эффектов цАМФ реализуется на молекулярном, клеточном и организменном уровнях. Хотя функции цГМФ исследованы мало, полагают, что этот пуриновый нуклеотид является антагонистом
цАМФ, выполняющего роль универсального регулятора репликации, транскрипции, трансляции, активности многих ферментов и экстрацеллюлярного передатчика («сигнал»-молекула) специфических стимулов и информации от одних типов клеток к другим. Считают, что, если цАМФ является стоп-сигналом пролиферации и одновременно пусковым сигналом дифференцировки, то цГМФ тормозит эти процессы. Существенное значение определенного уровня цАМФ в сохранении специфической дифференцировки было показано на различных опухолевых и трансформированных клетках, которые восстанавливали исходную дифференцировку при увеличении концентрации цАМФ внутри клетки. В то же время неопластическая трансформация всегда сопровождалась статистически достоверным снижением содержания цАМФ. На ts-мутантах онковирусов выявлена прямая корреляция между внутриклеточным уровнем цАМФ и некоторыми свойствами трансформированных клеток, такими как адгезивность, морфология, нечувствительность к межклеточным контактам, увеличение агглютинируемости под действием растительных лектинов и др. Более злокачественные клетки характеризуются низким содержанием цАМФ и большей чувствительностью к воздействию экзогенного цАМФ или же веществам, индуцирующим его внутриклеточный синтез. Добавление дибутирил-цАМФ и экзогенного цАМФ ингибирует рост злокачественно трансформированных клеток in vivo и in vitro, а также восстанавливает их морфологические и функциональные свойства (Р. Е. Кавецкий, 1977). Вместе с тем во многих быстрорастущих неоплазмах, наоборот, иногда определяется высокое содержание цАМФ и при повышенном уровне цАМФ стимулируется синтез ДНК. Вероятно, такие противоречивые данные можно объяснить сложностью взаимосвязей в этой регуляторной системе клетки. Очевидно, цАМФ активирует фосфорилирование регуляторных и структурных клеточных белков. Предполагают, что морфологическая нормализация трансформированных клеток цАМФ обусловлена его действием на активность поверхностных гликозилтрансфераз. Таким образом, большая совокупность признаков воздействия цАМФ, характеризующих трансформированный фенотип клеток in vitro, находится под контролем цАМФ. Получены менее убедительные данные, свидетельствующие о влиянии цАМФ на процессы дифференцировки, так как наблюдались и повышение, и снижение уровня дифференцировки при воздействии цАМФ на клетки.

Аденозин-3', 5'-циклофосфат. Этот нуклеотид
Нуклеозидтрифосфаты (Nucleoside triphosphate, NTP) — это нуклеозиды с тремя фосфатами. Природные нуклеозидтрифосфаты представлены аденозинтрифосфатом(ATP), гуанозинтрифосфатом (GTP), цитидинтрифосфатом (CTP), тимидинтрифосфатом (TTP) и уридинтрифосфатом (UTP). Данные термины означают, что нуклеотиды содержат сахар рибозу.
Нуклеотиды, содержащие сахар дезоксирибозу, имеют приставку дезокси- в имени и d- в сокращении: дезоксиаденозинтрифосфатом (dATP), дезоксигуанозинтрифосфатом (dGTP), дезоксицитидинтрифосфатом (dCTP), дезокситимидинтрифосфатом (dTTP) и дезоксиуридинтрифосфатом (dUTP).
К фосфатным остаткам нуклеотидов могут присоединяться один или два остатка фосфорной кислоты с образованием, соответственно, нуклеозиддифосфатов (НДФ) или нуклеозид — трифосфатов (НТФ).
На основе рибонуклеотидов образуются следующие рибонуклеозиддифосфаты и рибонуклеозидтрифосфаты:
Аденозиндифосфат (АДФ) и аденозинтрифосфат (АТФ); гуанозиндифосфат (ГДФ) и гуанозинтрифосфат (ГТФ); цитидиндифосфат (ЦДФ) и цитидинтрифосфат (ЦТФ); уридиндифосфат (УДФ) и уридинтрифосфат (УТФ).
Из дезоксирибонуклеотидов образуются дезоксирибонукле-озиддифосфаты и дезоксирибонуклеозидтрифосфаты:
Дезоксиаденозиндифосфат (дАДФ) и дезоксиаденозинтри-фосфат (дАТФ);
Дезоксигуанозиндифосфат (дГДФ) и дезоксигуанозинтри-фосфат (дГТФ);
Дезоксицитидиндифосфат (дЦДФ) и дезоксицитидинтри-фосфат (дЦТФ);
Дезокситимидиндифосфат (дТДФ) и дезокситимидинтри-фосфат (дТТФ).
Нуклеозиддифосфаты и нуклеозидтрифосфаты являются производными нуклеотидов. В них остатки фосфорной кислоты соединены между собой связями, обладающими большим запасом энергии. Такие связи принято называть макроэргическими (высокоэнергетическими) и обозначать значком а молекулы, содержащие такие связи, — макроэргическими соединениями.
Среди всех НДФ и НТФ особое место занимает аденозинтрифосфорная кислота (АТФ):
В ходе реакции гидролиза макроэргической фосфатной связи в молекуле АТФ происходит значительное уменьшение свободной энергии, составляющее около 7 ккал/моль. Эту энергию клетка может использовать для выполнения какой-либо химической работы. Для сравнения при гидролизе немакроэргической фосфатной связи уменьшение свободной энергии составляет приблизительно 2—3 ккал/моль.
АТФ является первичным аккумулятором энергии в клетке.
Энергия, необходимая для «зарядки» АТФ, поставляется в процессах диссимиляции — дыхания и брожения, а у зеленых растений — также в процессе фотосинтеза. В ходе диссимиляции в макроэргических связях АТФ запасается энергия химических связей распадающихся веществ, а при фотосинтезе — непосредственно световая энергия.
АТФ расходует энергию на процессы ассимиляции — различные биосинтезы, а также на транспорт веществ, совершение механической работы. Другие НТФ также участвуют в процессах ассимиляции. Например, УТФ обеспечивает энергией процесс биосинтеза сахарозы в растениях, ГТФ — клетчатки, ЦТФ — фосфолипидов. Однако все они являются вторичными аккумуляторами энергии в клетке, т. к. получают энергию от АТФ.
Таким образом, АТФ является не только первичным аккумулятором, но и универсальным источником энергии в клетке.
Это — энергетическая «валюта» клетки.
Фосфодиэфирная связь — высокоэнергетическая совокупность ковалентных связей, образуемая атомом фосфора в фосфатной группе и двумя молекулами посредством двух эфирных связей. Фосфодиэфирные связи играют ключевую роль во всех биологических системах, образуя остов нуклеиновых кислот ДНК и РНК.