82.Мембранні фосфо та гліколіпіди. Похідні сфінгозину.
Фосфоліпіди — похідні гліцеролу чи сфінгозину та жирних кислот Окрім цих сполук до складу фосфоліпідів входить фосфат (залишок фосфорної кислоти) і додаткові гідрофільні радикали, що формують полярну голівку молекули (аміноспирти — холін, етаноламін; амінокислоти — серин та інші). Фосфоліпіди чи гліцерофосфати — похідні гліцеролу (гліцерину) складаються з гліцерину, у якого перша і друга гідроксильні групи заміщені жирними кислотами, а третя — заміщена фосфатною групою, до якої приєднаний гідрофільний радикал. В результаті утворюється амфіфільна молекула (від грецького — та, що любить двох) — гідрофобні подвійні хвости, утворені вуглеводневими кінцями жирних кислот і гідрофільна компактна голівка, до складу якої входить фосфат і певний гідрофільний радикал. Голівка несе електричні заряди (+N; -0; 0; -СОО), але в більшості випадків нейтральні. Лише невелика кількість заряджені. Наприклад фосфатид серину має негативний заряд.
В склад гліцерофосфату входять насичені (стеаринова, пальмітинова та ін.) і не насичені (олеїнова, лінолева) висококарбонові жирні кислоти, число атомів вуглецю в яких варіює від 14 до 24. Насичені жирні кислоти розміщуються паралельно, не насичені — під кутом до осі молекули. Часто в складі гліцерофосфатів одна молекула жирної кислоти насичена, а інша не насичена. Остання також робить вигин в ділянці подвійного зв'язку, що забезпечує рухомість ліпідного шару.
Фосфоліпіди — похідні сфінгозину (фосфосфінголіпіди) складаються із сфінгозину — довгого аміноспирту СН3-(СН2)і2-СН=СН-СНОН-СНМН2-СН2ОН, у якого водень аміногрупи заміщений ліпідами або висококарбоновою жирною кислотою (церамід), а гідроксил — фосфатною групою, до якої приєднаний як і у гліцерофосфатидів, певний гідрофільний радикал (холін, серин, етаноламін і т. п.). Вуглеводневий радикал сфінгозину і жирної кислоти утворює 2 гідрофобних хвоста, тому фосфосфінголіпід також амфіфільний.
Гліколіпіди також можуть бути похідними гліцеролу (у прокаріот і рослин) чи сфінгозину (у тварин). Вони також мають гідрофобні вуглеводневі ланцюги (за рахунок двох жирних кислот глікогліцероліпідів чи один з ланцюгів від сфінгозинів, інший від жирної кислоти у глікосфінголіпідів) і полярну гідрофільну голівку. Остання утворена різноманітними вуглеводами приєднаними до останнього гідроксилу гліцерину чи сфінгозину.
83.Характеристика фаз мітозу.
Цитологічний аналіз фаз мітозу. Власне мітоз поділяють на 4 основні фази: П, М, А, Т.
Профаза: 1) Конденсація хромосом (третій рівень). 2) Регресія ядерця. Ядерцевий організатор інактивується, а матеріал ядерця диспергується: більша його частина йде в цитоплазму, інша, менша частина, перерозподіляється між ЯО. 3) В кінці П відбувається швидкий розпад ядерної оболонки на мілкі вакуолі, що не відрізняються від везикул ЕС. Ці фрагменти залишаються асоційованими з мітотичним веретеном. 4) Одна з основних подій П — побудова мітотичного веретена — головного компонента мітотичного апарату. Мікротрубочки двох сусідніх мітотичних центрів (у тваринних астральних центрів) сходяться утворюючи біполярне мітотичне веретено (складається з двох напівверетен). Мікротрубочки, що його утворюють називають полюсними. Після розпаду ядерної оболонки частина полюсних мікротрубочок вступає в контакт кінетохорами, що є начебто початком наступної фази мітозу.
Метафаза. 1) Посилення конденсації хромосом (4 рівень конденсації хромосом, хроматину). В цей період вона досягає максимуму, в результаті чого хромосоми стають найбільш короткими та товстими. Повністю відокремлюються одна від одної хроматиди, які утримуються разом в ділянці центромери. 2) Завершується формування веретена поділу, його асоціація з кінетохорами хромосом, рух хромосом. 3) Орієнтація хромосом на екваторі веретена у вигляді метафазної пластинки. Хромосоми орієнтуються по відношенню до осі веретена таким чином, що їх один полярний кінетохор обернений до одного із полюсів клітини, іншим — до другого полюсу і розміщуються в екваторіальній площині веретена, утворюючи фігуру у вигляді метафазної пластинки. До хромосом, розміщених в екваторіальній пластинці прикладені дві урівноважені сили спрямовані в протилежні сторони.
Анафаза. В анафазі одночасно зв'язок між хроматидами хромосом зупиняється, уже вільні сестринські хромосоми, колишні хроматиди відходять до протилежних полюсів зі швидкістю 1 мкм/хв. Розходження хроматид в анафазі відбувається в результаті двох незалежних процесів, що відбуваються в веретені: 1) деконденсація кінетохорних мікротрубочок та пов'язані з ними хроматиди за допомогою рухових білків відходять до полюсів; 2) подовження та ковзання мікротрубочок у кожному напівверетені за допомогою білка динеїна, що відбувається трохи пізніше, відштовхують полюса в протилежних напрямках.
Телофаза. В телофазі відбуваються процеси зворотні профазі. Хромосоми у полюсів фрагментарно деконденсуються, потоншуються і подовжуються. У місці контакту хромосом з мембранними бульбашками починається будуватись ядерна оболонка; одночасно полімеризуються білки трьох видів ляміни, що вистилають внутрішню ядерну мембрану, утворюються порові комплекси. Фрагменти ядерної оболонки на початку оточують окремі хромосоми, далі зливаючись, утворюють ядерну оболонку. Перш за все, хромосоми контактують з ядерною оболонкою центромерними і теломерними гетерохроматиновими ділянками. В пізній телофазі активується ЯО і починає утворюватись ядерце.
Цитокінез. Після телофази відбувається цитокінез. Перпендикулярно осі веретена в ділянці його екватора з внутрішньої сторони цитоплазматичної мембрани утворюється скорочувальне кільце. Воно починає утворюватись ще на початку анафази, але починає функціонувати після завершення телофази. Скорочувальне кільце в основному складається з активних компонентів, що знаходяться у комплексі з невеликою кількістю міозинових молекул. Механізм його скорочення ідентичний скороченню по типу ковзання міофібрил — актинових філаментів відносно міозинових філаментів за допомогою міозинових голівок. В міру скорочення кільця, поглиблюється борозна поділу, але саме кільце не потовщується, зберігається постійна товщина за рахунок видалення частини філаментів. Після завершення цитокінезу скорочувальне кільце повністю розпадається. В рослинних клітинах, що мають щільні клітинні стінки, цитокінез відбувається іншим способом — цитоплазма ділиться шляхом утворення нової клітинної стінки між двома дочірніми клітинами.
Окрім ядра, під час мітозу в цитоплазмі також відбуваються значні зміни.Мембрани ендоплазматичної сітки розпадаються на розрізнені мембранні вакуолі, апарат Гольджі розпадається на окремі диктіосоми. Деполімеризуються цитоплазматичні мікротрубочки і актинові філаменти. Зменшується кількість мітохондрій, пластид, лізосом. До метафази всі органоїди і структури відокремлюються веретеном до периферії клітини, а при цитокінезі йде пасивний їх розподіл по дочірніх клітинах.
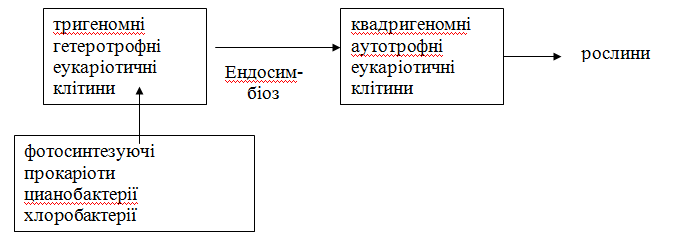
84.Які існують докази єндосимбіотичної теорії еволюції рослин грибів тварин. Складіть гіпотетичну схему, що включає: прокаріотичну екологічну систему; тригеномну еукаріотичну клітину; квадригеномну еукаріотичну клітину. При упорядкуванні схеми використовуйте такий гіпотетичний (проте який має вагомі докази) ланцюг еволюційних подій. У середині Архею метаболічне різноманіття досягло великої досконалості у всіх областях біосинтезу, яке могло б виникнути в елементарній клітинній системі: клітини могли синтезувати або утилізувати майже всі складні органічні речовини. Наслідком цього метаболічного різноманіття явилося виникнення екосистем: продуценти, консументи декількох порядків, редуценти. Проте прокаріотична екосистема виявилася тупиковою гілкою еволюції, тому що кожна прокаріотична клітина була вже метаболічно вузькоспеціалізованою. Еволюційний стрибок був пов'язаний із серією послідовних ендосимбіозів з утворенням багатофункціональної еукаріотичної системи.
Тригеномна еукаріотична клітина - прообраз клітин тварин і грибів - виникла з 3-х ендосимбіонтів. Нуклеоцитоплазма утворилася з великих анаеробних мікроорганізмів - хазяїв, але спроможних витримати високі температури і кислотність завдяки тому, що їх ДНК була захищена білками-гістонами. З ними ввійшли в ендосимбіоз (проникли усередину) аеробні мікроорганізми, згодом перетворившись у мітохондрії, що призвело до ефективності енергетичного обміну і синтезу нових складних органічних речовин, зокрема стероїдів, із якими пов'язане удосконалення багатьох метаболічних процесів: синтез гормонів, ліпоїдів, спроможність до піно- та фагоцитозу.
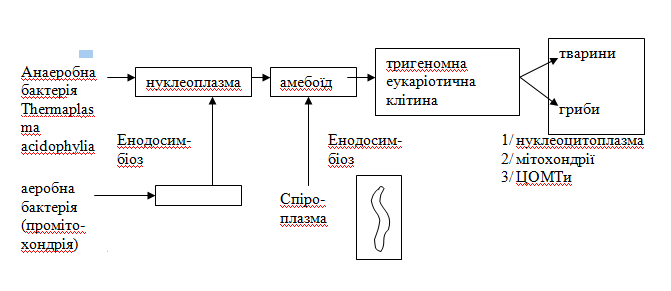
Наступним еволюційним кроком до еукаріотичної клітини було набування внутрішньо- і позаклітинної рухливості: центрів організаторів мікротрубочок. Їх привнесли прокаріотичні мікроорганізми за типом спіроплазм, що мають внутрішньотрубочкову систему.
Найостанніша еволюційна подія в серії послідовних ендосимбіозів - набуття еукаріотами спроможності до фотосинтезу. Для цього гетеротрофні еукаріоти фагоцитували фотосинтезуючих прокаріот, але не перетравлювали їх остаточно. Останні надалі еволюціонували в пластиди.