

Учебные материалы занятия
Клеточный цикл. В процессе жизненного цикла клетка удваивает свое содержимое и делится на две дочерние. В организме млекопитающего для поддержания жизни производятся ежесекундно миллионы новых клеток. Нарушение регуляции новообразования и размножения клеток проявляется как онкологическое заболевание. Этим вызван большой интерес к изучению и моделированию механизмов регуляции клеточного деления.
Схема клеточного цикла изображена на рисунок 1. Клеточный цикл состоит из двух периодов: митоз (М-фаза) включает разделение предварительно удвоенного ядерного материала, деление ядра, деление самой клетки, цитокинез и занимает около часа. Значительно более длительный период между двумя митозами занимает интерфаза, включающая стадию роста G1, фазу репликации ДНК (S), фазу подготовки к делению G2. Клеточный цикл (рис.2) регулируется генами и белками-ферментами двух основных классов. |
Рис. 1. Схема стадий клеточного цикла |

Ферменты циклин-зависимые протеинкиназы (Cdk) индуцируют последовательность процессов путем фосфорилирования отдельных белков – циклинов. Циклины – семейство белков-активаторов циклин-зависимых протеинкиназ (Cdk) – ключевых ферментов, участвующих в регуляции клеточного цикла эукариот.
Циклины, синтезируются и деградируют при каждом новом цикле деления, связываются с молекулами Cdk и контролируют их способность к фосфорилированию, без циклина Cdk не активны.
Циклины |
Циклин-зависимые протеинкиназы |
Фаза клеточного цикла |
Циклин Е |
Cdk 2 |
Вход в S-фазу |
Циклин А |
Cdk 2 |
Вход в G2-фазу |
Циклин В |
Cdk 1 |
Вход в М-фазу |
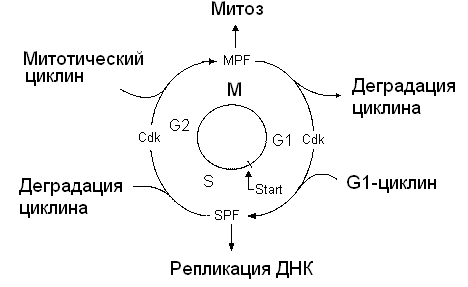
Рис. 2. Схема регуляции клеточного цикла
Количество этих молекул регуляторов в клетках организмов различно. Например, в делении дрожжевой клетки принимают участие один Cdk и девять циклинов, образующие девять разных циклин-Cdk комплексов. У более сложно организованных млекопитающих изучено шесть Cdk и полтора десятка циклинов. Контроль выхода клетки из G1-фазы и G2-фазы осуществляют промотор-фактор S-фазы (S-PF) и промотор-фактор M-фазы (M-PF). Они представляют собой гетеродимеры. Cуществует особая контрольная точка клеточного цикла (Start), с которой заканчивается рост (G1-фаза) и начинается процесс синтеза ДНК.
Регуляция клеточного цикла. Регуляция клеточного цикла осуществляется в процессе обратимого фосфорилирования/дефосфорилирования регуляторных белков. Ключевым белком, регулирующим вступление клетки в митоз (G2/M-переход), является специфическая серин/треонин-протеинкиназа, которая носит название фактор созревания – ФС (начало митоза – M-PF). В активной форме фермент катализирует фосфорилирование многих белков, принимающих участие в митозе, например, факторы транскрипции, белки митотического веретена и ряд ферментов. Фосфорилирование этих белков запускает процесс митоза. Фосфорилирование белков является важным регулятором их ферментативной активности. Фосфорилирование существенно меняет химические свойства белков. В результате белок становится способным распознать, связать, активировать, деактивировать, фосфорилировать или дефосфорилировать свои субстраты. Таким образом, фосфорилирование может включать и выключать ферменты.
После завершения митоза регуляторная субъединица ФС, циклин, маркируется белком убиквитином и подвергается протеолизу, т.е. процессу гидролиза пептидных связей. Теперь наступает очередь протеинфосфатаз, которые дефосфорилируют белки, принимавшие участие в митозе, после чего клетка возвращается в состояние интерфазы.
ФС – фермент, включающий регуляторную субъединицу, циклин и каталитическую субъединицу, циклинзависимую киназу. Активной формой фермента является лишь димер ЦЗК + циклин. Кроме того, активность протеинкиназы регулируется путем обратимого фосфорилирования самого фермента.
В клетках позвоночных присутствует ряд различных циклинов и циклинзависимых киназ. Разнообразные сочетания двух субъединиц фермента регулируют запуск митоза, начало процесса транскрипции в G1-фазе, начало процесса репликации ДНК в S-периоде интерфазы (стартовый переход) и другие ключевые переходы клеточного цикла.
В ооцитах лягушки вступление в митоз (G2/M-переход) регулируется путем изменения концентрации циклина. Циклин непрерывно синтезируется в интерфазе до достижения максимальной концентрации в фазе М, когда запускается весь каскад фосфорилирования белков, катализируемый ФС. К окончанию митоза циклин быстро разрушается протеиназами, также активируемыми ФС. В других клеточных системах активность ФС регулируется за счет различной степени фосфорилирования самого фермента.


Примечания:
Хроматида – это одна нить ДНК или молекула ДНК;
хромосома – структура, состоящая из 2-х сестринских хроматид.
В S-периоде клеточного цикла происходит синтез ДНК, то есть для каждой имеющейся молекулы ДНК синтезируется одна копия (сестринская ДНК). Обе молекулы, старая и вновь синтезированная, частично соединены между собой, и в профазу митоза или мейоза из таких двух молекул ДНК (хроматид) образуется одна хромосома.
Всегда следует помнить, что перед тем как клетка вступает в митоз или мейоз она проходит G1-, S-, G2- периоды клеточного цикла. В S-периоде происходит синтез или редупликация ДНК. В результате этого количество молекул ДНК удваивается.
Митоз
Продолжительность митоза зависит от размеров клеток, их плоидности, числа ядер, а также от условий окружающей среды, в частности от температуры. В животных клетках митоз длится 30-60 мин, в растительных 2-3 часа. Более длительны стадии митоза, связанные с процессами синтеза; самодвижение хромосом осуществляется быстро.
Регуляция митоза. В организме митоз контролируются системой нейрогуморальной регуляции, которая осуществляется нервной системой, гормонами надпочечников, гипофиза, щитовидной и половых желёз, а также местными факторами (продукты тканевого распада, функциональная активность клеток). Взаимодействие различных регуляторных механизмов обеспечивает как общие, так и местные изменения митотической активности. Митоз опухолевых клеток выходят из-под контроля нейрогуморальной регуляции.
Выражением регуляции митоза в связи с взаимодействием организма и среды служит суточный ритм деления клеток. В большинстве органов ночных животных максимум митоза отмечается утром, а минимум – в ночное время. У дневных животных и человека отмечается обратная динамика суточного ритма. Суточный ритм митоза – следствие цепной реакции, в которую вовлекаются ритмические изменения внешней среды (освещённость, температура, режим питания и др.) и биологические ритмы (ритм функциональной активности клеток и изменения процессов обмена веществ).
Нарушения митоза. При различных патологических процессах нормальное течение митоза нарушается. Выделяют 3 основных вида патологии митоза:
1) повреждения хромосом (набухание, склеивание, фрагментация, образование мостов, повреждения центромеров, отставание отдельных хромосом при движении, нарушение их спирализации и деспирализации, раннее разъединение хроматид, образование микроядер;
2) повреждения митотического аппарата (задержка митоза в метафазе, многополюсный, моноцентрический и асимметричный митоз, трёхгрупповая и полая метафазы). Особое значение в этой группе патологии митоза имеет колхициновый митоз, или К-митоз, который вызывается алкалоидом колхицином и другими так называемые статмокинетическими ядами, используемыми в качестве мутагенов. К-митозы возникают и самопроизвольно в культуре ткани и опухолях. При К-митозе нарушаются расхождение центриолей и поляризация ими веретена деления, подвергается дезорганизации митотический аппарат, не происходит разъединения хроматид (К-пары).
3) Нарушения цитотомии, т.е. разделение тела растительной или животной клетки. Патологические митозы возникают после воздействия митотических ядов, токсинов, экстремальных факторов (ионизирующее излучение, аноксия, т.е. отсутствие кислорода в организме или в отдельных органах, тканях, гипотермия), при вирусной инфекции и в опухоли. Резкое увеличение числа патологических митозов типично для злокачественных опухолей.
Мейоз
Особенности мейоза:
1) происходит только в половых клетках;
2) состоит из двух последовательных делений с короткой интерфазой между ними;
3) профаза первого деления очень сложная и отстоит из 5 стадий;
4) в зиготену, вторую стадию профазы редукционного деления, происходит соединение гомологичных хромосом (биваленты или тетрады), которые остаются связанными между собой до анафазы 1-го деления;
5) в анафазу 1-го деления происходит разделение и расхождение к полюсам целых хромосом, состоящих из двух хроматид, а не разделение хромосом на отдельные хроматиды, как в митозе;
6) в интерфазу между 1-м и 2-м делениями мейоза отсутствует S-период и перед 2-м делением не происходит редупликации ДНК;
7) в процессе мейоза образующиеся дочерние клетки полностью не разделяются между собой, а остаются связанными тонкими цитоплазматическими мостиками.
Продолжительность мейоза. Продолжительность мейоза напрямую зависит от количества ДНК в ядре. Также продолжительность мейоза зависит от структуры хромосомной организации и особенностей данного вида.
Вид |
2n |
Время, ч |
ДНК на клетку (пг) |
Львиный зев Antirrhinum majus |
16 |
24.0 |
5.5 |
Гаплопаппус изящный Haploppapus gracilis |
4 |
36.0 |
5.5 |
Рожь посевная Secale cereale |
14 |
51.2 |
28.7 |
Лук репчатый Allium cepa |
16 |
96.0 |
54.0 |
Традесканция палудоза Tradescantia paludosa |
12 |
126.0 |
59.0 |
Тульбагия фиолетовая Tulbaghia violacea |
12 |
130.0 |
58.5 |
Лилия Генри Lilium henryi |
24 |
170.0 |
100.0 |
Лилия длинноцветковая Lilium longiflorum |
24 |
192.0 |
106.0 |
Триллиум прямостоящий Trillium erectum |
10 |
274.0 |
120.0 |