

§ 9. Электронный микроскоп
Одним из важнейших итогов развития геометрической электронной оптики явилось создание электронного микроскопа. Этот прибор имеет такое же назначение, как и его оптический аналог: он служит для получения увеличенных изображений предметов, невидимых невооруженным глазом. Однако, если для обычного микроскопа ограничения, связанные с разрешающей силой и обусловленные дифракционными эффектами, не позволяют рассматривать объекты с размерами, меньшими 2000 — 3000 А, то применение электронного микроскопа довело предел разрешения до нескольких ангстрем. Завоевание этих трех порядков величины имело огромное значение; в результате оказалось возможным прямое наблюдение таких интересных объектов, как вирусы, коллоидные частицы, молекулы больших размеров.
Причина того, что современный электронный микроскоп по разрешающей силе настолько превосходит оптический инструмент, связана с различием в длине волны электронных и световых лучей.
В отсутствие иммерсии минимальное разрешаемое микроскопом расстояние между самосветящимися точками объекта определяется по формуле
![]() (9.1)
(9.1)
где
![]() /2
- апертурный угол (рис. 9.1). Если в микроскоп
рассматривают освещенные, а не
самосветящиеся объекты, т.е. отдельные
точки объекта рассеивают падающие на
них волны, исходящие из одной и той же
точки источника, и, следовательно,
свет, идущий из разных точек объекта,
оказывается когерентным, то формула
(9.1) принимает вид
/2
- апертурный угол (рис. 9.1). Если в микроскоп
рассматривают освещенные, а не
самосветящиеся объекты, т.е. отдельные
точки объекта рассеивают падающие на
них волны, исходящие из одной и той же
точки источника, и, следовательно,
свет, идущий из разных точек объекта,
оказывается когерентным, то формула
(9.1) принимает вид
![]() (9.1a)
(9.1a)
Как видим, различие совершенно не существенно. У современных микроскопов sin( /2) доходит до 0,95, так что возможно разрешение деталей, имеющих размеры около половины длины волны света.
Для электронных лучей длина волны де-Бройля определяется формулой
![]() (9.2)
(9.2)

Рис. 9.1. Апертурный угол.
Здесь U - разность
потенциалов, пройденная электронами,
формирующими изображение. Формула
(9.2) написана с учетом релятивистских
поправок. Для сравнительно медленных
электронов можно пользоваться приближенной
формулой
![]() ,
которая для U < 50 кB дает ошибку, не
превышающую 2%. Величина U
в этой формуле выражена в вольтах, а
,
которая для U < 50 кB дает ошибку, не
превышающую 2%. Величина U
в этой формуле выражена в вольтах, а
![]() - в ангстремах. Если принять U
= 100 кB (обычное значение напряжения в
электронном микроскопе), то
= 0,04А. Если бы в электронном микроскопе
можно было использовать такие же большие
апертуры, как в обычной оптике, то
разрешающая сила прибора достигала бы
десятых или даже сотых долей ангстрема.
Однако, как уже говорилось ранее, в
электронной оптике гораздо труднее
устранить оптические ошибки. В результате
в электронной микроскопии приходится
пользоваться пучками с небольшой
апертурой. Важнейшей оптической ошибкой,
ухудшающей качество изображения в
электронном микроскопе, является
сферическая аберрация, растущая
пропорционально кубу апертурного угла
.
Уменьшая угол
,
можно, в принципе, довести сферическую
аберрацию до сколь угодно малой величины,
но при этом будет возрастать дифракционное
размытие
- в ангстремах. Если принять U
= 100 кB (обычное значение напряжения в
электронном микроскопе), то
= 0,04А. Если бы в электронном микроскопе
можно было использовать такие же большие
апертуры, как в обычной оптике, то
разрешающая сила прибора достигала бы
десятых или даже сотых долей ангстрема.
Однако, как уже говорилось ранее, в
электронной оптике гораздо труднее
устранить оптические ошибки. В результате
в электронной микроскопии приходится
пользоваться пучками с небольшой
апертурой. Важнейшей оптической ошибкой,
ухудшающей качество изображения в
электронном микроскопе, является
сферическая аберрация, растущая
пропорционально кубу апертурного угла
.
Уменьшая угол
,
можно, в принципе, довести сферическую
аберрацию до сколь угодно малой величины,
но при этом будет возрастать дифракционное
размытие
![]() .
Очевидно, что наивыгоднейшее значение
будет примерно соответствовать условию
равенства размытий, вызываемых обоими
эффектами. При выполнении этого требования
оказывается, что апертурный угол должен
быть порядка 1/1000 радиана, а разрешающая
сила микроскопа, которая определяется
как оптическими ошибками и дифракционным
размытием, так и рядом трудно устранимых
дефектов, обусловленных несовершенством
конструкции электронно-оптической
системы, составляет около 2-3 А.
.
Очевидно, что наивыгоднейшее значение
будет примерно соответствовать условию
равенства размытий, вызываемых обоими
эффектами. При выполнении этого требования
оказывается, что апертурный угол должен
быть порядка 1/1000 радиана, а разрешающая
сила микроскопа, которая определяется
как оптическими ошибками и дифракционным
размытием, так и рядом трудно устранимых
дефектов, обусловленных несовершенством
конструкции электронно-оптической
системы, составляет около 2-3 А.
В основных чертах конструкция электронного микроскопа, предназначенного для получения изображения тонких объектов в проходящих электронных лучах, повторяет конструкцию оптического микроскопа. Главными элементами конструкции электронного микроскопа просвечивающего типа являются: 1) источник электронов — электронная пушка; 2) конденсорная линза, с помощью которой электронный пучок фокусируется на объект; 3) объектив и 4) проекционная линза.
Ход лучей в микроскопе рассматриваемого типа с магнитными линзами показан на рис. 9.2. В целях наглядности на том же рисунке изображена схема обычного микроскопа. Объектив формирует промежуточное, действительное и увеличенное изображение рассматриваемого предмета в плоскости АВ. Проекционная линза дает окончательное изображение на флюоресцирующем экране или на фотографической пластинке в плоскости А1В1. Для получения большого общего увеличения объектив и проекционная линза должны обладать маленьким фокусным расстоянием. Технически короткофокусные линзы легче осуществить в магнитном варианте. Главным недостатком короткофокусных электростатических линз является то, что для фокусировки электронного пучка с энергией 100 кэB требуется применение очень сильных электрических полей. Последнее обстоятельство приводит к ряду трудностей в связи с появлением нежелательных утечек и пробоев. Сказанное не означает, однако, что электростатические электронные микроскопы не используются. Существует ряд систем с линзами этого рода.

Рис. 9.2. Схематическое изображение оптического микроскопа и электронного (магнитного) микроскопа просвечивающего типа.
Поясним процесс образования изображения при просвечивании объекта в электронном микроскопе. Использование простой оптической аналогии здесь исключено, так как механизм возникновения изображения в обоих случаях совершенно различен.
В оптическом микроскопе контраст между различными элементами изображения обусловлен различным поглощением световых лучей в соответствующих точках объекта. В электронном микроскопе поглощение электронов в объекте практически не происходит (сильное поглощение разрушило бы объект!), но электроны испытывают рассеяние, т. е. меняют направление своего движения. При этом участки объекта, обладающие большей плотностью или большей толщиной, сильнее рассеивают проходящие пучки. Различие в степени рассеяния и используется для получения контрастного изображения объекта.
Используемый метод несколько различается в зависимости от того, какое изображение требуется получить: светлопольное или темнопольное. Остановимся сначала на первом случае.

Рис. 9.3. Формирование светлопольного изображения.

Рис. 9.4. Формирование темнопольного изображения. Осветительная система перекошена.
Вблизи объектива, на главной оптической оси системы, помещается апертурная диафрагма с маленьким отверстием. Такая диафрагма пропускает только те электроны, которые при прохождении через объект меняют направление своего движения лишь на очень маленький угол, и срезает все лучи, отклоненные на большие углы (рис. 9.3). В результате менее плотные или более тонкие участки объекта будут выглядеть на изображении более светлыми.
В случае темнопольного изображения лучи, испытавшие малое рассеяние, с помощью того или иного искусственного приема исключаются из процесса формирования изображения. Поэтому более тонкие участки объекта окажутся на изображении более темными. Один из наиболее удобных приемов, позволяющих осуществить эту процедуру, состоит в перекашивании осветительной системы микроскопа (рис. 9.4).
Темнопольное изображение обладает определенным преимуществом перед светлопольным — оно позволяет получать более контрастные снимки объекта. Поясним сказанное примером.
Пусть некоторый участок объекта, размещенный на подложке (например, на тонкой бесструктурной коллодиевой пленке), рассеивает вне апертурного угла около 10% проходящего электронного потока, а сама подложка — около 5%. Тогда в случае светлопольного изображения отношение электронных освещенностей будет составлять 95:90, т. е. контрастность будет явно недостаточной. В случае же темнопольного изображения соответствующее отношение освещенностей составит 1:2 и контрастность будет хорошей.
Еще один вопрос требует разъяснения. В случае оптического микроскопа селективная по длинам волн поглощательная способность различных участков объекта отвечает окрашенному предмету и, следовательно, цветному изображению. Применение красителей, которые неодинаково связываются различными участками рассматриваемой структуры, позволяет резко повысить контрастность бледного изображения объекта. В электронном микроскопе изображение на экране монохроматично, но возможность искусственного повышения контраста остается и в этом случае. Роль «красителей» здесь играют атомы тяжелых элементов, на которых происходит более сильное рассеяние электронного пучка. Если удается обеспечить избирательную сорбцию таких атомов на определенных участках рассматриваемой структуры, то эти детали оказываются выделенными на изображении.
Электронная микроскопия превратилась в обширную, самостоятельную, быстро развивающуюся область современной технической физики. Открытие новых, неизвестных ранее ультраструктур, установление новых неожиданных пространственных соотношений произошло в результате применения электронного микроскопа в физике, химии и кристаллографии, но нигде оно не носило столь революционного характера, как в биологии. Электронный микроскоп позволил вести наблюдение фагов и привел к установлению их структуры, привел к открытию неведомых деталей внутреннего строения клеток, в частности рибосом (центров белкового синтеза), позволил, наконец, осуществить прямое наблюдение молекул ДНК - носителей генетической информации организмов. Решение центральной проблемы молекулярной биологии - установление связи между структурой и функцией - представляется теперь более близким.
В качестве примеров, иллюстрирующих сказанное, приведем несколько снимков биологических объектов, полученных с помощью электронного микроскопа.

Рис. 9.5а. Электронный снимок фага Т-2 с увеличением в 180000 раз.
На рис. 9.5а приведен снимок фага Т-2 с увеличением примерно в 180000 раз. Как видно из снимка, частица фага состоит из головки, ограниченной плоскими гранями, и хвостового отростка белковой структуры (см, рис. 9.5 б). Только в результате выяснения строения фаговой частицы удалось понять механизм взаимодействия фага с клеткой. Адсорбция фага на поверхности клеточной мембраны приводит к химической реакции, в ходе которой осуществляется проникновение в клетку инфекционной части вирусной частицы — нуклеиновой кислоты. Белковая оболочка фага при этом остается снаружи, как это подтверждается прямыми электронно-микроскопическими наблюдениями. Проникнув в клетку, молекула ДНК фага, с одной стороны, навязывает клетке процессы белкового синтеза, необходимые для роста вируса, а с другой, сама начинает с большой скоростью реплицироваться. В результате уже через короткий промежуток времени (~103 сек) внутри клетки образуются сотни новых вирусных частиц. Параллельно с размножением фага в клетке происходят глубокие изменения: клетка перестает делиться и вскоре погибает вследствие разрушения клеточной оболочки новыми фаговыми частицами, которые входят в среду и могут заражать новые клетки.

Рис. 9.56. Схема структуры фага Т-2.
На рис. 9.6 а приведены электронно-микроскопические снимки, а на рис. 9.6 б — схема процесса взаимодействия вируса с клеткой.
Электронный снимок рибосом представлен на рис. 9.7 а, их схематическое изображение - на рис. 9.7 б. Наблюдаемые только в электронном микроскопе рибосомы имеют молекулярные размеры, их поперечник порядка 100—150 А. Эта внутриклеточная структура состоит из двух субъединиц.
Заметим, что выяснение строения рибосом имеет фундаментальное значение для раскрытия механизма белкового синтеза.

Рис. 9.6 а. Электронный снимок бактериальной клетки. Слева - начало атаки вируса, справа — клеточная оболочка разрушена.

Рис. 9.6 б. Схематическое изображение процессов, видимых на снимках предыдущего рисунка.
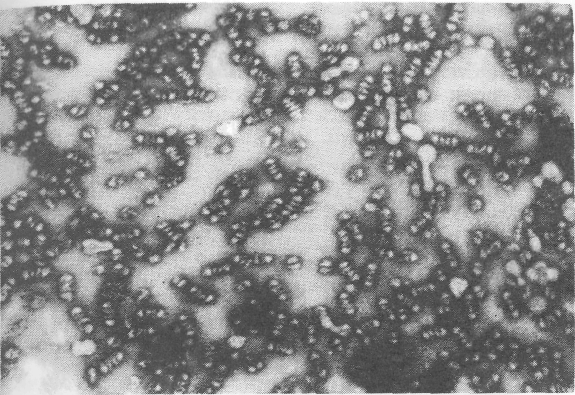
Рис. 9.7'а. Электронный снимок рибосом. Увеличение х 130000.
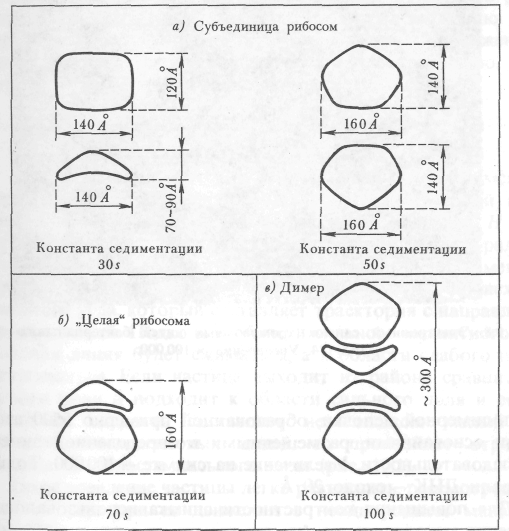
Рис. 9.76. Схематическое изображение рибосом.
Еще один пример: на рис. 9.8 представлен снимок трех кольцевых хромосомных нитей бактериального вируса ФХ-174.
В сущности, каждая хромосома в данном случае представляет собой макромолекулу ДНК, двойная спираль которой состоит из полимерной цепочки, образованной примерно 5500 азотистыми основаниями, размещенными в определенной линейной последовательности. Увеличение на снимке - 100000. Толщина спирали ДНК - около 20 А.
Для повышения контрастности снимка использовалось напыление металла.

Рис. 9.8. Электронный снимок хромосомных нитей бактериального вируса ФХ-174. Увеличение х 100000.