

1ivanov_a_a_fiziologiya_ryb
.pdfОднако о причинах смещенной активности ведутся споры. Тинберген видел причину в конфликте двух биологических мотиваций, имеющих исключительно большое значение. Избыточная энергия первой мотивации (спаривание) разряжается за счет второй (ухаживание за гнездом).
Подобным образом разрешаются и конфликтные ситуации. Вблизи построенного гнезда (своя территория) самец колюшки активно нападает на чужака, вторгшегося на его территорию. Попав на чужую территорию, тот же самец не нападает на хозяина, а активно ретируется на свою территорию.
Однако, если самцы встречаются на границе владений, конфликт разрешается иначе. Они проявляют смещенную активность. На границе участков самцы принимают позы копательного поведения, что с точки зрения целесообразности лишено всякого смысла (рис. 15.24).
В данной ситуации две сильные мотивации (атаковать или бежать?) разряжаются во внешне бессмысленном копательном поведении.
Не совсем понятен и механизм выбора приоритетности мотиваций. Одновременно в организме рыбы формируется несколько потребностей и, следовательно, несколько мотиваций. В течение одного дня животные осуществляют различные виды полезного поведения. В более или менее константных условиях жизни у животного складывается определенный распорядок, в соответствии с которым каждый мотивированный потребностью вид деятельности занимает определенное время суток. Остается неясным, каким образом происходит распределение времени по видам активности. Заслуживает специального изучения и механизм перераспределения времени по видам активности в случае появления дополнительного сильного биологически значимого фактора.
Рис. 15.24. Копательная поза (смешенное поведение) агрессивных самцов, встретившихся на границе своих территорий
Д. Мак-Фарленд (1988) ввел в этологию животных понятие упругости поведения, которое несколько проясняет ситуацию.
Упругость поведения - это мера того, до какой степени может быть сжат во времени любой вид активности животного под влиянием других видов. Данное явление характерно для всех животных независимо от уровня их эволюционного положения. Рыбы не являются исключением из этого правила. Время, затрачиваемое, например, парой лялиусов на исследовательскую деятельность, пищевую активность и игровое поведение, перераспределяется по мере созревания самки. Чем ближе момент нереста, тем меньше времени уделяется игровому поведению. В определенный момент в поведении самца появляются новые виды активности: ухаживание за самкой и построение гнезда. Параллельно ограничивается время на другие виды активности. После нереста самец сокращает время на питание, исследовательскую активность, игры до нуля. Весь суточный запас времени в этот период занимает активность, связанная с ухаживанием и охраной гнезда. В других ситуациях более
эластичным оказывается пищевое поведение.
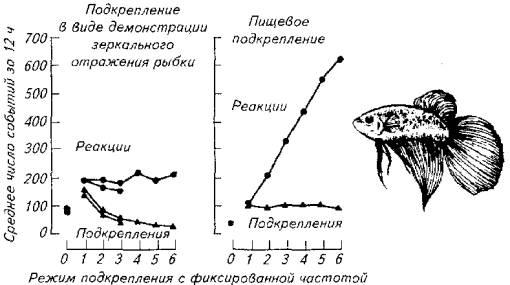
Эластичность поведения иллюстрирует и следующий опыт на бойцовых рыбках
(рис. 15.25).
У бойцовой рыбки вырабатывали условный рефлекс на прохождение через специальный тоннель, помещенный в аквариум.
Рис. 15.25. Эластичность агрессивного поведения (слева) и эластичность пищевого поведения у бойцовой рыбки
Появление рыбки в тоннеле подкрепляли зеркальным отражением той же особи, чем провоцировали агрессивность самца. В другом варианте опыта использовали пищевое подкрепление.
Как следует из диаграммы ответов, агрессивная активность оказалась неэластичной
вотличие от пищевой активности рыб. В другом варианте время, отводимое на пищевое поведение, расширялось по мере увеличения частоты подкрепления ответных реакций. Количество проходов через тоннель в течение 12ч наблюдений при подкреплении зеркальным отражением не зависело от частоты предъявляемого подкрепления.
По принципу функциональной системы осуществляется не только регуляция поведения рыб (набор моторных реакций), но и поддержание гомеостаза. Константы гомеостаза делят на жесткие и мягкие (изменяющиеся в более широких пределах). Для рыб жесткими константами гомеостаза являются уровень кислорода в крови, осмотическое давление внутренней среды. В широких же пределах изменяются температура тела, давление крови, уровень глюкозы в крови, концентрация сывороточных белков и жирных кислот в крови.
В качестве примера рассмотрим функциональную систему постоянства кислорода в крови. Это гомеостатическая функциональная система с активным внешним звеном саморегуляции.
Результатом действия данной системы является поддержание кислорода на определенном уровне. Его изменения возбуждает (угнетает) дыхательный центр (группа нейронов спинного, продолговатого, промежуточного мозга). Возникающее возбуждение поступает в гипоталамус, где в результате афферентного синтеза формируется программа нормализации уровня кислорода в крови, которая включает
всебя соматический (поведенческий) и вегетативный механизмы. Они запускаются напрямую из гипоталамуса или через влияния по нисходящим нервным путям (рис. 15.26).
Соматический ответ заключается в том, что рыба ищет воду с высоким содержанием
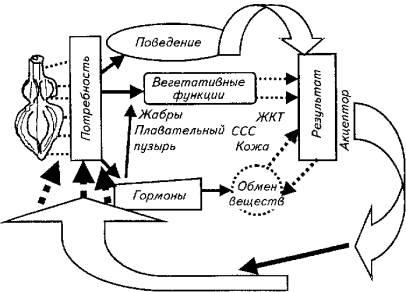
кислорода или поднимается к поверхности и жадно хватает атмосферный воздух. Вегетативная регуляция сводится к целому спектру влияний на жаберный аппарат (повышаются частота дыхания, проницаемость через жаберные мембраны кислорода), сердечно-сосудистую систему (возрастает частота сердечных сокращений, выбрасываются в русло депонированные эритроциты). Происходят адаптационные изменения в метаболизме рыбы, включается гормональная регуляция вспомогательных вегетативных процессов. Дополнительно кислород перекачивается из плавательного пузыря, включаются в газообмен кожа и желудочно-кишечный тракт.
И соматические, и вегетативные влияния приводят к одному результату: количество поступающего в организм рыбы кислорода возрастает, его потребление снижается. Акцепторы результата информируют гипоталамус о произошедших изменениях.
Если полезный приспособительный эффект достигнут, регулируемая функция гипоталамуса тормозится. Если результат не удовлетворяет лимбическую систему и поставленная цель не достигнута, процесс соматовегетативной регуляции повторяется.
Рис. 15.26. Функциональная система поддержания количества кислорода в крови рыб Обратная связь осуществляется посредством гуморального канала (О2, СО2, рН-
рецепторы гипоталамуса оценивают состав крови непосредственно) и нервной афферентации от соответствующих рецепторов периферических органов. Таким образом, данная функциональная система хотя и относится к гомеостатическому типу, но включает все возможные эффекторные рычаги регуляции - поведенческий, вегетативный, метаболический.
В заключение хочется еще раз подчеркнуть, что теория функциональных систем дает возможность исследователю объяснить цельное поведение рыбы независимо от того, сколь сложна биологическая задача (потребность).
Эта теория применяется для объяснения не только поведения отдельных индивидуумов, но и для трактовки стайного (группового) поведения рыб. Формирование групповых отношений определяется врожденными причинами, которые закрепляют биологическую целесообразность группового образа жизни. Так, трудно представить стайный образ жизни крупных хищников. С другой стороны, мало шансов на выживание в одиночестве у верховки, уклеи. Изоляция от группы уклеи приводит ее в дискомфортное, стрессовое состояние, из которого рыба старается выйти как можно быстрее, объединившись со стаей.
Встае существуют сложные иерархические отношения; каждая особь в стае занимает определенное место и свою специфическую групповую функцию. Правда, эта позиция индивидуума не прочна; она постоянно подвергается переоценке. Каждый член стаи старается поднять свой статус и занять более выгодное положение.
Структура стаи у разных видов рыб может быть различной, Однако в стае с лидером есть позиции более и менее выгодные, что и побуждает особей к постоянным иерархическим отношениям. Так, менее выгодно положение в конце стаи: последним меньше достается корма, а в случае преследования они первые кандидаты в жертвы хищника. Объединение в стаю выгодно ее членам: в стае им комфортно и безопасно (менее опасно).
Всвою очередь, стая полезна для популяции. Стая удовлетворяет СБОИ пищевые, оборонительные, репродуктивные функции и потребности популяции вида в целом. Стая значительно эффективнее адаптируется к изменениям среды обитания, так как,
содной стороны, индивидуальный положительный опыт становится достоянием всех членов стаи, а с другой - стая обеспечивает проверенным временем полезным групповым опытом каждого члена стаи.
Вприроде постоянно происходит некоторое перемешивание групп, особенно заметное при миграциях молоди. Последнее также полезно для популяции, так как сводит к минимуму вероятность инбридинга. Кроме того, стая стабильна тогда, когда в ней достаточное количество особей.
Однако полезным результатом функциональной системы группового поведения все равно является равновесное состояние стаи. Стая тем стабильнее, чем большее количество ее членов стереотипично и синхронно реагируют на дестабилизирующий стаю фактор.
Функциональная система поведения стаи складывается из нескольких биологически детерминированных поведенческих блоков:
системы пищевого поведения; системы оборонительного поведения; системы игрового поведения; системы полового поведения; системы ориентировочного поведения,
Структурированная стая наиболее эффективно достигает полезноприспособительного результата.
Таким образом, как индивидуальное, так и стайное поведение рыб нацелено на удовлетворение определенных биологических потребностей. Использование стайного или индивидуального образа жизни рыбами - это всего лишь способ решения определенной биологической задачи.