

30 Сучасні методи ідентифікації бактерій.
1.Микроскопические методы включают приготовление мазков и препаратов для микроскопирования. В большинстве случаев результаты микроскопических исследований носят ориентировочный характер, так как многие микроорганизмы лишены морфологических и тинкториальных особенностей. 2.Микробиологические методы позволяет точно установить факт наличия возбудителя в исследуемом материале: включает культивирование, выделение чистой культуры и идентификацию микроорганизмов с учетом морфологических, тинкториальных, культуральных, биохимических, токсигенных и антигенных свойств. 3. Биологические методы направлены на определение наличия токсинов возбудителя в исследуемом материале и на обнаружение возбудителя, включают заражение лабораторных животных с последующим исследованием их. 4. Серологические методы выявления специфических антител и антигенов возбудителя - важный инструмент в диагностике инфекционных заболеваний. 5. Аллергологические методы. Антигены многих возбудителей обладают сенсибилизирующим действием, что используют для диагностики инфекционных заболеваний (кожно-аллергические пробы).
31 Цисты и споры
это специализированные клеточные структуры бактерий. Многие бактерии способны образовывать структуры, помогающие им переживатьт в течение длительного времени неблагоприятные условия и переходить в активные вегетативные формы при попадании в подходящие для этого условия. Цисты встречаються у различных групп бактерий: азотобактера, спирохет, миксобактерий, риккетсий. У большинства миксобактерий образование цист – закономерная стадия их жизненного цикла. После окончания стадии активного розмножения клетки миксобактерий собираются в месте и образуют так называемые плодовые тела. Внутри плодовых тел клетки переходят в покоящее состояние. В одних случаях покоящие клетки могут морфологически не отличаться от вегетативных, в других их образование сопровождается заметными структурными и морфологическими изменениями. Такие изменение называют микроцистами. Они функцыонально аналогичны бактериальным эндоспорам: более устойчивы к нагреванию, высушиванию, розличным физическим воздествиям, чем вегетативные клетки.
Эндоспоры. Эндоспоры образуются бактериями, принадлежащими к родам Bacillus, Clostridium, Sporosarcina и некоторым другим. Бактерии образуют српоры, когда создаются такие условия в окружающей среде, которые индуцируют процесс спорообразования. Считается, что споры не обя зательная стадия цикла развития спорообразующих бактерий.
Факторы, индуцирующие спорообразование,различны: недостаток питательных веществ в среде, изменение pH, температуры, накопление выше опредиленного уровня продуктов клеточного метобализма. Для бактериальных эндоспор характерна устойчивость к высокой температуре, воздействиям различных токсических веществ, радиации и другим неблагоприятным факторам. Это связывают прежде всего с ттем, что содиржимое споры находится в сильно обезвоженном состоянии.
В каждой бактериальной клетке, как правило, формируется одна спора.
Формирование споры начинается с того, что в зоне локализации ядерных нитей происходит уплотнение цитоплазмы, которая в месте с генетичеким материалом обособляется от остального клеточного содержимого с помощью перегородки. Следующий этап формирования споры – обрастание отсеченного участка клеточной цитоплазмы с ядерным материалом мембранной вегетативной клетки, конечным результатом которого является образование проспоры – структуры, расположенной внутри материнской клетки и полностью отделенной от нее двумя елементарными мембранами: наружной и внутренней по отношению к проспоре. Эти этапы формирование споры обратимы.
Процесс спорообразования протекает под контролем генетического аппарата материнской клетки, в генетическом материале которой есть участки, ответственные за отдельные этапы процесса.
Сформированные бактериальные эндоспоры могут находится в жизнеспособногм состоянии в течение длительного времени, При создании благоприятных условий такие споры проростают и дают начало вегетативной клетке, проростание – это процесс, сопровождающийся сложными физиологическими и биохимическими изменениями. Начинается оно с интенсивного поглощения спорой воды и набухания. На первом этапе проростания происходит активация ферментов, резко возрастает дыхание. Последующие этапы состоят в розрушении кортекса, розрыве споровой оболочки, выходе из нее сформировавшейся к этому времени структуры, называемой ростовой трубочкой, достраивании ею клеточной стенки и последующим делении сформированной вегетативной клетки
32 Механізм субстратного фосфорилювання.
Реакции, в которых энергия, освобождающаяся на определенных окислительных этапах брожения запасается в молекулах АТФ , получили название субстратного фосфорилирования . К синтезу АТФ по механизму фосфатного фосфорилирования ведут катаболические реакции, которые в зависимости от своей химической природы могут быть разделены на 2 типа. Большинство относится окислительно-восстановительным реакциям. Богатые энергией соединения возникают в процессе брожения на этапах анаэробного окисления. Например, окисление фосфоглицеринового альдегида ( ФГА), катализирующие ФГА- дегидрогеназой, приводит к образованию богатого энергией метаболита -1,3- дифосфоглицериновой килоты. Второй тип реакций связан с расщеплением субстратов или промежуточных продуктов, образующихся из них. Катализируются эти реакции ферментами, относящимися к классу лиаз. Например, у гетероферментативных молочнокислых бактерий высокоэнергетический ацетилфосфат обр. из ксилулозо-5-фосфата в реакции, катализируемой фосфокетолазой:
Кси-5-фосфат + Фп→ФГА+ ацеилфосфат + Н2О
Из других высокоэнергетических соединений важное место в энергетике процессов брожения принадлежит фосфоенолпировиноградной кислоте(ФЕП). Эти соединения харак. Тем что свободная энергия,освобождающаяся при их гидролизе находится в области -35 до -88 кДж/моль и с помощью соответствующих ферментов может быть перенесена на молекулы АДФ.
Все р-ии суб. фосфорилирования локализованы в цитозоле клетки, что указывает на простоту хим. Механизмов в основе суб. фосфорилирования.
33 Особливості біосинтезу мікробною клітиною полісахаридів та ліпідів.
Липиды – группа, в которой обединены различные по своей природе соединения жиры сложные эфиры глицерина и жирных кислот), высокомолекулярные жирные кислоты, воска( сложные эфиры высших жирных кислот и спиртов), каратиноиды, а также некоторые витамины. Общими свойствами липидов являются их нерастворимость в воде и растворимость в органических растворителях. У прокариотов липиды входят в состав клеточных мембран и клеточной стенки, а также служит запасными веществами клетки.
Синтез жирных кислот с четным числом углеродных атомов происходит в результате последовательного присоединения к молекуле ацетил – КоА, двухуглеродного остатка. Сначала из ацетил – КоА образуется малонил – КоА. Пути, ведущие к синтезу малонил – КоА различные. Один из них состоит в карбоксилировании метильной группы ацетил – КоА, ферментом является ацетил – КоА- карбоксилазой:
СН3 – СО S КоА + СО2 + АТФ → НООС – СН2 – СО S – КоА + АДФ + Фн
В последующих реакциях конденсации карбоксильная группа снова отщепляется в виде СО 2. Синтез жирных кислот происходит при участии мультиферментного комплекса согласно уравнению:
Ацетил – КоА + 7 Малонил – КоА + 14 NADPH2 → Пальметил – КоА + 14 NADP + 7 СО2 + 7 КоА + 7 Н2О.
Полисахариды.
Микроорганизмы способны синтезировать моно, олиго и полисахариды, а также другие соединения, в состав которых входят углеводы. Если в культуральной среде присутствует глюкоза то она может использываться для синтеза компонентов клетки, являющихся ее производными. Гетеротрофы обладают способностью синтезировать глюкозу из С 2 и С 3 – соединений, а автотрофы из СО2.
У автотрофов Глю из углекислоты синтезируется в цыкле Кальвина, основной реакцией которого является образование из СО 2 и рибулозо – 1,5 – дифосфата 3 фосфоглицериновой кислоты; из нее уже путем преобразований получается Глю. При этом процессе также как и при биосинтезе глюкозы, у гетеротрофов используются в основном реакции протикающие при катаболизме глюкозы в гликолитическом пути, но идущие в обратном направлении.
У прокариотов широко развита способность к взаимопревращению сахаров. Как правило осуществляется взаимопревращение не свободных сахаров, а их нуклеозиддифосфопроизводных. Например, глюкозо – 1 – фосфат может реагировать с УТФ, образуя УДФ глюкозу с высвобождением неорганического пирофосфата. УДФ глюкоза может потвергаться метаболическим превращением с образованием ряда других сахаров. Другие нуклеозиды также могут выступать в реакциях взаимопревращений и образовывать нуклеозиддифосфосахара.
34
35 . Гіпотеза Мітчела. Механізми окислювального та фотосинтетичного фосфорилювання.
П. Митчелл – английский биохимик, который высказал гипотезу, получившую название хемоосмотической или электрохимической. Согласно этой гипотезе необходимой промежуточной стадией на пути трансформации энергии окисления в химическую энергию макроэргических фосфатных связей является электрическая энергия. Основой постулат гипотезы Митчелла состоит в том что « дыхание и фосфорилирование связаны между собой черезе электрохимический потенциал ионов водорода на мембране.» Предполагается, что перенос водорода по дыхательной цепи осуществляется определенно ориентерованными ферментами перпендикулярно мембране. Сами мембраны, несущие ферменты электронного транспорта и сопряженного сним фосфорилирования, в интанктном состоянии непроницаемым для протонов и ионов. Но легко проницаемым для электронов.
Механизм окислительного фосфорилирования
Отданные субстратами восстановительные эквиваленты (протоны и элетроны) переносятся на плазматическую мембрану. Через мембрану они траснпортируются таким образом, что между внутренней и внешней сторонами мембраны создается элетрохимический градиент с положительным потенциалом снаружи и положительным внутри. Это перепад заряда возникает благодаря определенному расположению компонентов дыхательной цепи в мембране.
Некоторые из этих компонентов переносят элетроны, другие переносят водород, Взаиморасположение переносчиков таково, что при транспорте элетрона от субстрата к кислороду протоны (Н+) связываются на внутренней стороне мембраны, а освобождаются на внешней. Эта система траспортирующая элетроны и протоны получила название дыхательной или электроно-транспортной цепи.
Мембрана содержит специальный фермент АТР- синтазу, синтезирующий АТР из АДП и Р. Этот фермент выступает из мембраны с ее внутренней стороны. В процессе синтеза АТР за счет энергии транспорта элетронов через мембрану называют окислительным фосфорилированием или фосфорилированием в дыхательной цепи.
Фотосинтетическое фосорилирование
Фотосинтетическое фосфорилирование и образование НАДФ 2 приисходит у цианобактерий следующим образом: квант света выбивает элетрон из рекационного центра фотосистемы I и переносит его на переносчик Х, от которого электрон далее попадает на НАДФ+ с образованием НАДФН 2. Для использования элетронов воды у цыанобактерий синтезируются специаьные фотохимически реакционные центры, включающие хлорофилл и фикобилипротеиды. Окислительно-востановительный потенциал содержащего эти ферменты фотореакционого центра фотосистемы II более положительный чем потенциал воды. Еще один квант света выбивает электрон из фотосистемы II и переносит его к преносчику У на более высокий энергетический уровень,чем в фотосистемы I. Таким образом он заполняет свободное место в фотосистеме I. После нужно заполнить такое же место и в фотосистеми II. Для этого используется электрон оторванный от воды. При переносе электрона от переносчика У до реакционного центра в фотосистеме I энергия электрона используется в реакции АДФ+Ф= АТФ. Это фосфорилирование обеспечивает серия переносчиков, распалагающихся в порядке понижения их окислительно – восстановительного потенциала.
36 Нуклеиновые кислоты (НК)– это линейные неразветвленные гетерополимеры, мономерами которых являются нуклеотиды, связанные фосфодиэфирными связями.
2 типа нуклеиновых кислот: ДНК и РНК. НК обеспечивают хранение, воспроизведение и реализацию генетической (наследственной) информации. Нуклеотиды – это органические вещества, молекулы которых состоят из остатка пентозы (рибозы или дезоксирибозы), к которому ковалентно присоединены остаток фосфорной кислоты и азотистое основание. Азотистые основания в составе нуклеотидов делятся на две группы: пуриновые (аденин (А) и гуанин(Г)) и пиримидиновые (цитозин(Ц), тимин(Т) и урацил(У)). Дезоксирибонуклеотиды включают в свой состав дезоксирибозу и одно из азотистых оснований: А, Г, Т, Ц. Рибонуклеотиды включают в свой состав рибозу и одно из азотистых оснований: А,Г,У,Ц. ДНК – это НК, мономер - дезоксирибонуклеотиды. ДНК является первичным носителем наследственной информации. Обычно молекула ДНК прокариот - кольцевая двойная правозакрученная спираль, которая состоит из двух нитей (или цепей), связанных между собой водородными связями. Каждая нить - чередующиеся остатки дезоксирибозы и фосфорной кислоты, причем, к дезоксирибозе ковалентно присоединяется азотистое основание. При этом азотистые основания двух нитей ДНК направлены друг к другу и за счет образования водородных связей образуют комплементарные пары: А=Т (две водородных связи) и Г≡Ц (три водородных связи). Поэтому нуклеотидные последовательности этих цепей однозначно соответствуют друг другу. РНК – это НК, мономер - рибонуклеотиды. Азотистые основания образуют пары А=У, А=Т и Г≡Ц. Благодаря этому возможна передача информации от ДНК к РНК, от РНК к ДНК и от РНК к белкам.
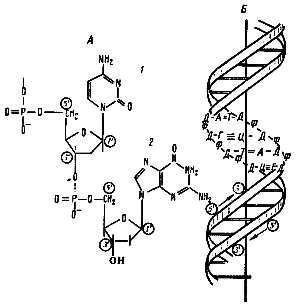
Строение ДНК: А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: 1 — цитозин; 2 — гуанин; Б — двойная спираль ДНК: Д — дезоксирибоза; Ф — фосфат;
37