Sos репарация у е. Сoli.
Данный тип репарации необходим для прохождения репликативной машиной повреждений. 1 этап – сигналинг о повреждении. RecA связывается с однонитевой ДНК и образует ДНК-белковые филаменты (однонитевые участки ДНК образуются при остановке репликативных вилок, так как полимераза3 перестает работать, а хеликаза дальше расплетает). В клетке есть много генов, которые в норме не работают (как раз и нужны для прохождения репликации через повреждения), иначе в норме в клетке будет много мутаций. LexA - регулятор транскрипции генов, кодирующих участвующие в репарации повреждений ДНК белки (димеры Lex A связываются с SOS боксами (20 п.н. консенсусы) в операторах генов репарации и ингибируют транскрипцию). Филаменты RecA стимулируют аутопротеолиз LexA, в результате чего происходит активация генов SOS-репарации.
UmuDC оперон – ген, кодирующий особую черезблоковую полимеразу – она может проходить через повреждения, включая при этом рандомно любой нуклеотид. После прохождения повреждения ее необходимо убрать и опять посадить нормальную репликативную полимеразу (иначе будет очень много мутаций из-за случайного включения нуклеотидов). Как именно работает этот самый оперон - UmuD повергается автопротеолитическому расщеплению с образованием активного фрагмента UmuD’. UmuD’ активирует черезблоковую полимеразу UmuC. Комплекс (UmuD’)2-UmuC теперь называют ДНК полимераза V (эта полимераза осуществляет репликацию через AP сайты, тимидиновые димеры и ряд других повреждений). Важно, чтобы эта полимераза работала недолго. Для работы черезблоковой полимеразы нужен ее контакт с PCNA и RecA-филаментом. При движении полимераза разрушает этот самый филамент и как только он разрушается полностью, черезблоковая полимераза сваливается и садится нормальная репликативная3 (скотосбрасыватель). Черезблоковая полимераза успевает синтезировать всего пару нуклеотидов. У каждого организма целый набор черезблоковых полимераз.
Репарация двунитевых разрывов.
Двунитевые разрывы в ДНК возникают под действием ионизирующего излучения, окислительного стресса,под действием некоторых химических агентов, в частности, ингибиторов ДНК топоизомеразы II (существуют в том числе природные ингибиторы). топоизомеразы нужны для релаксации ДНК, но если если действуют их ингибиторы, то остается двунитевой разрыв. С однонитевыми разрывами гораздо проще – они сшиваются лигазами. Существует два основных пути репарации двунитевых разрывов: гомологичная рекомбинация и негомологичное соединение концов ДНК.
Гомологичная рекомбинация: необходим донор гомологии (гомологичная хромосома – но это никогда у эукариот, или гомологичная хроматида – есть сразу после репликации).
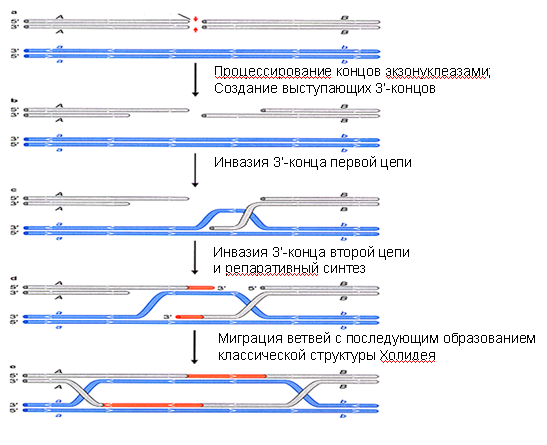
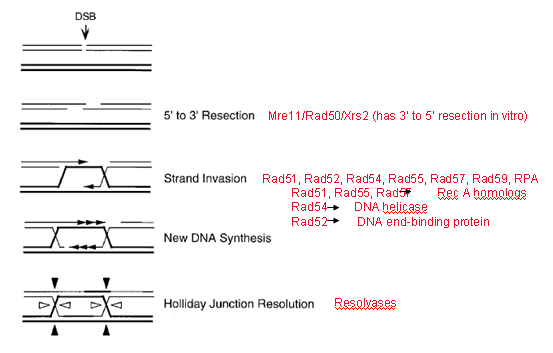
Экзонуклеазы создают свободный 3’-конец, который внедряется в соседний дуплекс, после чего происходит репликация с нормальной матрицы достаточно длинного фрагмента, затем второй переброс, в итоге образуется классическая структура Холидея.
Но нужна особая система внедрения, ведь ДНК стабильная (какой-то белок, облегчающий инвазию цепи). У E. Coli ключевым компонентом системы гомологичной рекомбинации
является белок RecA, который связывается с однонитевой ДНК и обеспечивает ее инвазию в двойную спираль с образованием D-петли.
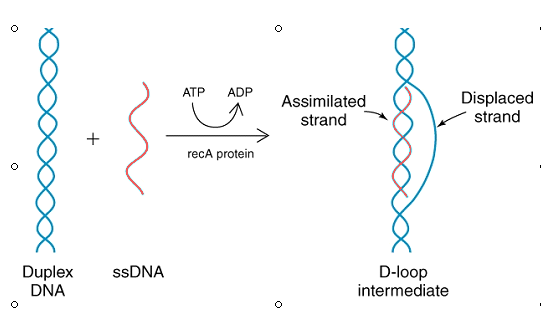
Это было выяснено в экспериментах, в которых смешивали одно- и двунитевую ДНК (они содержали какой-то участок гомологии), и ничего не происходило. А потом добавляли RecA и часть двойной спирали вытеснялась.
В эукариотических клетках гомологом recA является Rad51. Покрытая Rad51 однонитевая ДНК внедряется в гомологичный участок сестринской хроматиды с образованием D петли . 3’-конец внедрившейся цепи достраивается ДНК полимеразой и отжигается с 3’-концом комплементарной цепи исходного дуплекса. Бреши застраиваются и однонитевые разрывы лигируются. В итоге получаются две восстановленные ДНК.
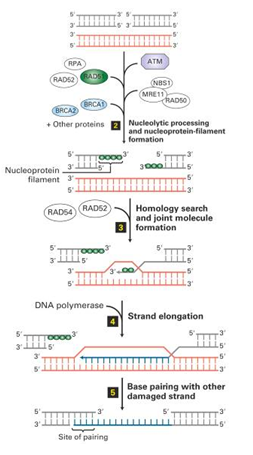
У эукариот есть альтернативный аналог этого прокариотического процесса, с полным восстановлением гомологии, без ошибок:
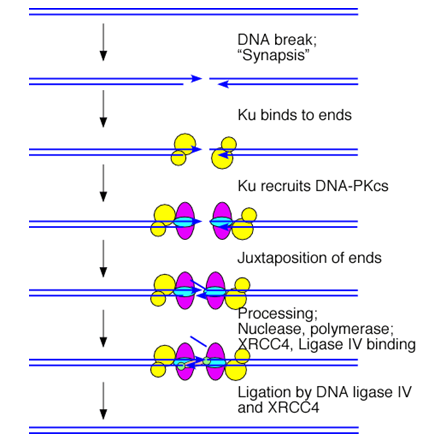
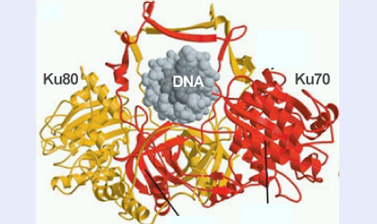
Но все-таки, гомологичная рекомбинация обычно чаще используется прокариотами, а вот эукариоты обычно лечат двунитевые разрывы негомологичным соединением концов – просто расчет на то, что свободные разорванные концы рядом. Ku (очередное кольцо) – заходит со стороны разрыва и удерживает ДНК, Далее оно привлекает все ферменты, в том числе ДНК-зависимые протеинкиназы. Далее происходит процессирование концов. Но точно воспроизвести исходную структуру не получается. Репликации здесь совсем нет. Концы делаются лигируемыми за счет утраты части информации (но эта информаия может быть и неважная). Но могут также быть и различные перестройки, в том числе приводящие к опухолям. В конце лигирование ДНК. Ku – это очередное кольцо, оно связывается и обхватывает ее, при этом удерживая, два витка вокруг ДНК, работает в виде димера (разрыв при этом доступен другим белкам).