

Нейротрансмиттеры первичных аффекторов
Приняты следующие критерии, позволяющие признать какое-либо соединение нейротрансмиттером первичных афферентных ноцицепторов:
наличие этого вещества в синапсах первичных аффекторов заднего рога;
высвобождение данного вещества при повреждающей стимуляции;
введение вещества должно давать такой же эффект, как и стимуляция первичных аффекторов;
назначение антагонистов блокирует как действие этого вещества, так и первичных аффекторов [2].
Многие вещества претендовали на роль нейротрансмиттеров первичных аффекторов ноцицепторов, например субстанция Р, соматостатин, сосудисто-активные интестинальные полипептиды. Однако ни одно из них не удовлетворило всем приведенным выше требованиям.
Субстанцию Р обнаруживают в пластинах I и II, соматостатин - в пластине II, а полипептиды - в пластине I. После пересечения заднего корешка содержание всех этих соединений в заднем роге быстро снижается [2]. Субстанция Р вызывает такую же деполимеризацию нейронов второго порядка в заднем роге, как и стимуляция первичных афферентов [70]. Однако ее нельзя признать только первичным афферентным нейротрансмиттером, поскольку она присутствовала менее чем в 25% клеток ганглия заднего корешка. Кроме того, ноцицепция сохранялась даже после разрушения субстанции Р [71]. Другие пептиды, в том числе холецистокинин, гастринвысвобождающий пептид, динорфин, энкефалин, ангиотензин II и бомбезин также обнаруживались в первичных афферентах и могли играть роль в ноцицепции.
Между тем продолжается накопление доказательств существования первичных афферентных нейротрансмиттеров. Доказано, что существуют быстрые и медленные компоненты, стимулирующие нейроны задних рогов. Субстанция Р напоминает медленный компонент. Аминокислоты, такие как глутаминовая и аспарагиповая, могут проводить быстрый компонент стимуляции.
Трансмиссионные клетки в спинном мозге Анатомия
В заднем роге содержатся клетки трех классов:
1) проекционные клетки, передающие информацию ростральным центрам;
2) возбуждающие промежуточные нейроны, обеспечивающие дальнейшую передачу к проекционным клеткам, к другим промежуточным нейронам или к двигательным клеткам, осуществляющим простые рефлексы;
3) угнетающие промежуточные нейроны, моделирующие ноцицептивную трансмиссию [74].
Критериями, позволяющими отличить проекционный нейрон, являются максимальная реакция при возбуждающей стимуляции, а также расположение нейрона в зоне, вовлеченной в поцицепцию. Нейроны, соответствующие этим критериям, концентрируются в пластинах I, II и V.
Большинство нейронов пластины I реагируют на повреждающее действие. В этой пластине происходит конвергенция многих афферентных нейронов. Поэтому рецептивные поля каждого из таких первичных афферентных нейронов становятся в несколько раз больше, чем его собственная зона рецепции. Клетки пластины I могут получать сигналы по миелинизированным и по немиелинизированным волокнам. Некоторые клетки возбуждаются только от поцицепторов (ноцицептивпо-специфичсские клетки [НС]). Значительное большинство клеток пластины I являются НС-клетками, или нейронами с высоким порогом возбуждения. Остальные клетки этой пластины получают дополнительную информацию от неноцицептивных аффекторов. Такие клетки получили названия нейронов широкого динамизма (НШД), поскольку они реагируют на широкий спектр стимулов разной интенсивности (рис. 2-29). Большинство клеток пластины 1 являются проекционными, передающими сигнал дальше в таламус.
В пластине II также содержится много ноцицептивных клеток. Большинство из них - это промежуточные нейроны, соединяющиеся с другими клетками этой пластины, расположенные в одном двух соседних сегментах. Многие клетки из других пластин связаны своими дендритами с клетками пластины II, а это подразумевает, что последняя может воздействовать на другие участки серого вещества спинного мозга. Так, клетки одного типа («стержневые» клетки) посылают сигналы в пластину I [75, 76]. Клетки другого типа («островные» клетки), по-видимому, являются ингибиторами, некоторые из них могут обладать разносторонним энергетическим влиянием [77-79].
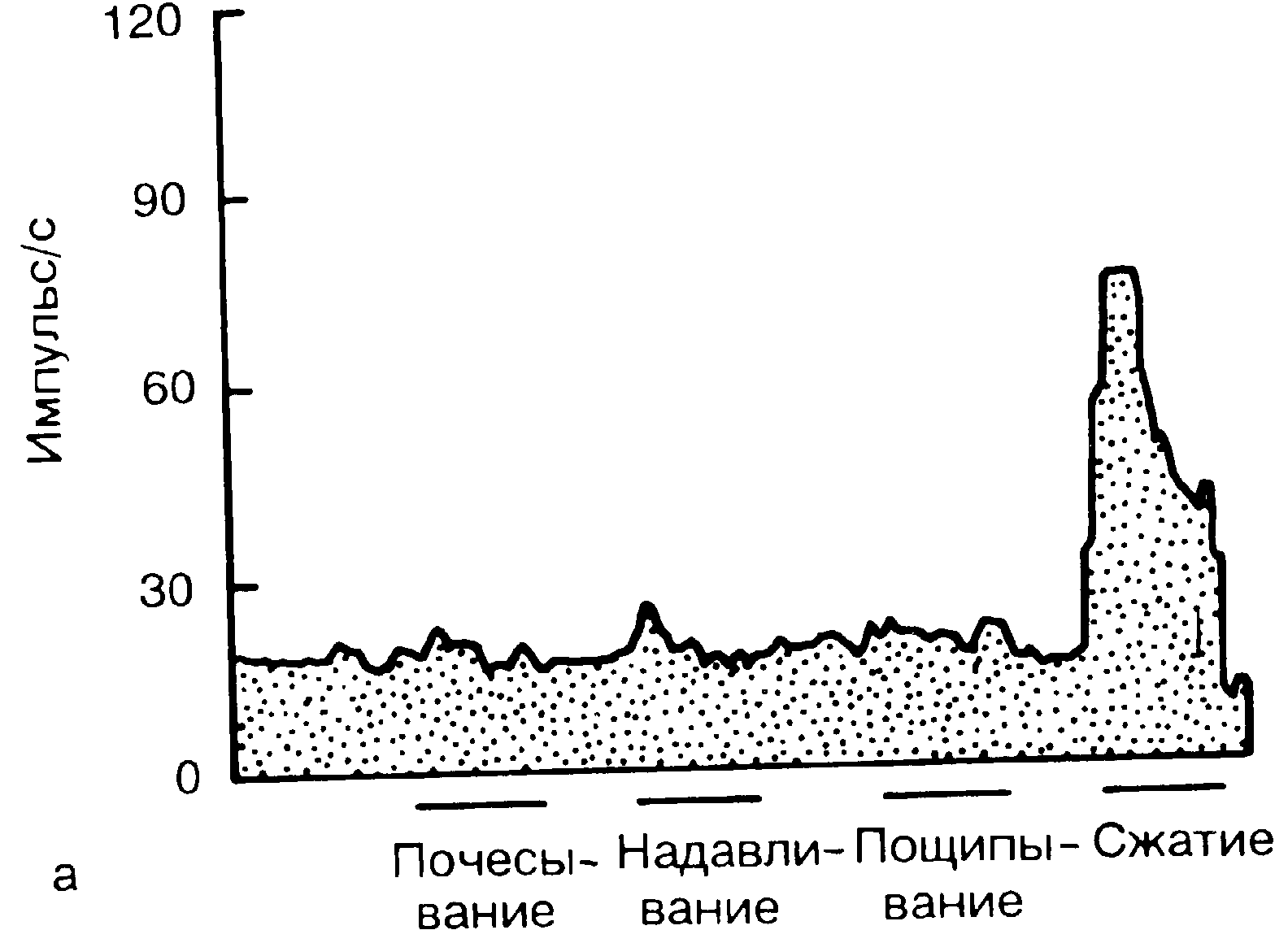

Рис. 2-29. Различие между ноцицсптивно-специфическими нейронами (а) и нейронами широкого динамизма (б) в заднем роге.
Ноцицептивно-специфические нейроны слабо реагируют на все безвредные стимулы (почесывание, надавливание, пощипывание) (а), но бурно отвечают на повреждающее воздействие, например сжатие (б). Нейроны широкого динамизма реагируют и на безвредные воздействия. Степень их реакции возрастает по мере усиления повреждающего воздействия. (Модифицировано по Fields [2].)
Нейроны пластин III и IV, как правило, очень восприимчивы к неповреждающим воздействиям и связаны синапсами преимущественно с большими миелинизированными афферентами. Некоторые из этих клеток связаны с другими пластинами и могут моделировать ноцицепцию, а другие дают отростки до таламуса. Пластина V, подобно пластине I, связана в основном с ноцицепцией. Ее клетки имеют обширные рецептивные поля благодаря конвергенции афферентных сигналов из разных источников. Дендриты клеток этой пластины проникают в слои пластин I и II. Афферентные сигналы из пластины I передаются промежуточными нейронами через пластину IV к пластине V. Большинство клеток последней относятся к типу НШД. В этой же пластине расположено большинство тел клеток спиноталамического тракта. Кроме того, значительное большинство клеток пластины V дают аксоны, доходящие до таламуса.
На ведущую роль пластин I, II и V в ростральной ноцицептивной трансмиссии указывают такие признаки, как:
клетки этих структур дают аксоны вплоть до таламуса;
пересечение этих путей устраняет восприятие боли с кожи [79 ];
раздражение, нанесенное в области пластин I и II, вызывает боль [79];
импульс в этих клетках нарастает по мере усиления раздражения [80];
кратковременное раздражение нейронов I и II пластин вызывает раннюю и позднюю реакции, аналогичные «первой» и «второй» боли [20, 81].
Пластина VI получает преимущественно сигналы проприорецептивного характера и проецирует их в мозжечок. В пластинах VII и VIII содержится много ноцицептивных нейронов, получающих сложные сигналы с обширных рецептивных полей [82]. Последующая передача этих сигналов происходит в таламус и в ретикулярную формацию.
Клетки пластины Х принимают в основном ноцицептивные сигналы с малых рецептивных полей (подобно клеткам I и II пластин) [83].
Суммируя сведения, приведенные выше, можно отметить, что пластины I, II и V являются основными зонами конвергенции ноцицептивной трансмиссии в спинном мозге. В пластинах I, V, VII и VIII содержится большая часть нейронов, обеспечивающих ростральную проекцию. Пластина II состоит преимущественно из промежуточных нейронов, пластина 1 - из ноцицептивно специфических клеток, а пластина V - из НШД. Нейроны пластин VII и VIII воспринимают сложные афферентные импульсы с обширных рецептивных полей.