

Тимин_О_А_Лекции_по_общей_биохимии-1
.pdfСтроениобменлипиедов |
|
|
|
|
|
|
|
|
|
250 |
|
|
|
|
|
|
|
|
|
||||
чествонеобразованныхФАДН |
|
|
2 соответчислудвойныхсвязейтвует. |
|
|
Остальныереа |
к- |
||||
циикладутбези менений. |
АТФ,потрнактивациюченной |
|
|
|
|
|
|
||||
o количествоэнергии |
|
|
|
. |
|
|
|
||||
|
|
Пример1Окис. пальмитиновойение |
|
|
кислоты |
|
|
||||
Таккакимеетсяатомов16угле, приода |
|
|
|
|
β-окисленииобразуется |
8молекулац |
е- |
||||
тил-SКоА .ПоследнийпоступаетвЦТК,приегоокисленводноборотецмиобразуеклаи |
|
|
|
|
|
|
т- |
||||
сямолекулы3 НАДН |
(7,5АТФ) |
,молекула1 ФАДН |
|
2 (1,5АТФ) |
имолекула1 ГТФ,чтоэкв |
|
и- |
||||
валентно1 |
0 молекуламАТФ. |
|
Итак, |
8молекулацетил |
|
-SКоА |
обеспечатобразование |
8×10=80 |
|||
молекулАТФ. |
|
|
|
числоциклов |
β-окисленияравно7 |
|
|
|
|||
Дляпальмитиновойкислоты |
|
.Вкаждомцикле |
|
б- |
|||||||
разуетсямолекула1 ФАДН |
онидадут"" |
2 (1,5АТФ) |
имолекула1 НАДН |
|
|
(2,5АТФ) |
.Поступаявдыхат |
|
ель- |
||
нуюцепь |
, всумме |
|
4 молекулы АТФ. Такимобразом, |
благодаря 7цикла |
м образу- |
||||||
ется 7×4=28 молекулАТФ. |
|
|
||
|
Двойныхсвязей |
впальмитиновойкислоте |
нет. |
|
|
Наактивациюжирнойкислоидемолекула1 тАТФы,которгидролизуется,одн, ако |
|
|
|
доАМФ,тоесть |
тратятсямакроэргические2 связи |
. |
|
|
|
Такимобразом,суммируя,получаем |
80+28–2 =106 молекул АТФ. |
||
|
Длярасчета |
коэффициента P/O (смОкислительное" фосфорилирование/Коэффициент |
||
P/O")подсчитываем |
|
|
||
o |
количествоАТФ,образованногомитохондрияхАТФ |
|
-синтазой – этос ответствует |
|
|
значению P, |
|
|
|
o |
общееколичествоНАДНФАДН |
2,поступившихдыхательнуюцепь |
– этозначение |
|
|
соответчислувосстановтвуетатомовкислорода( енных |
|
O),включенныхмолекулу |
|
|
воды. |
|
|
|
Послерасчетовполучаем |
P=72+28=100и |
O=32+14=46Такимобразом,величина. |
P/O |
||||
для β-окиспальмитиновойениякислоты |
равна2, |
17. |
|
|
|
||
|
|
Пример2Окисление. линолево |
|
йкислоты |
|
|
|
Т.к.числоатомовуглерравно18,т да |
|
количесмолекулацетилво |
|
-SКоА равно9 |
. |
||
Значит вЦТКобразуется |
9×10=90 молекулАТФ. |
|
|
|
|
||
Числоциклов |
β-окисленияравно8 |
.Прирасчетеполучаем8 |
×4=32 молекулы АТФ. |
|
|||
Вкислоте |
имеются 2двойныесвязи |
.Следовательно , вдвухциклах |
β-окисленияне |
б- |
|||
разуетсямолекулы2 ФАДН |
|
2,чторавнпотереценно |
|
3 молекулА |
ТФ. |
. |
|
Наактивациюжирнойкислоты |
тратятмакроэргические2 вязи |
|
|
||||
Такимобразом, |
энергетическийвыход |
окислениясоответствует |
|
90+32−3−2=117 моле- |
|||
кул АТФ. |
коэффициента P/O получаем P=81+32=113и |
|
|
|
|||
Прирасчете |
O=36+14=50Такимобр . |
а- |
|||||
зом,величина |
P/O для β-окисления линолевой кислоты равна2, |
26. |
|
|
|||
|
|
|
|
|
|
||
|
|
К Е Т ОТНЕ ОЛАВ Ы Е |
|
|
|
||
|
|
|
|
|
|||
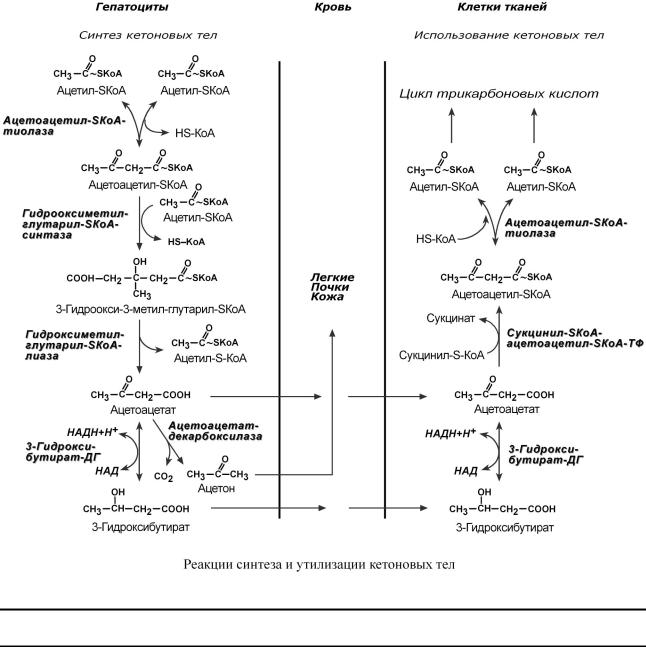
Ккетоновымтеламотносяттрисоединенияблизкойструктуры |
|
|
|
– ацетоацетат, |
|||
3-гидроксибутиратацетон. |
|
|
|
|
|
|
|
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
251 |
|||
|
|
|
|
|
||
|
Стимуломдляобразованиякетоновыхтелс поступлениеужитбольшогоколич |
|
|
е- |
||
стважирныхкислотв |
печень.Какужеуказыв,присостоянактивирующих, лось яхп |
|
о- |
|||
лизвжирткани,ов30%ообразованныхлойжирныхкислотзадерживапечень.К ются |
|
|
|
|||
этимсостояниямотнг ситсялодание, |
|
декомпенсация сахарного диабета I типа,длительные |
||||
физическиенагрузки,бо |
гатаяжирамидиета. |
Вобычныхусловияхсинтез |
|
кетоновых тел |
||
такжеидет,ховнаяменьшемногоколичестве. |
|
|
|
|
||
|
Удетейдолет7подвлияниемразличныхстикратк( голулов,инфекциидание, |
|
|
|
||
эмоциональноевозбуждение)ускоряетсясинкетонтелезмлегкожетвозникатьых |
|
|
е- |
|||
тоацид,сопронеукрзвождрвотацетонем("ющиойиморвота")йПр.сяичинойеская |
|
|
|
|||
этомуслуж |
итнеустойчуглеводобимвостьалыезапасынгликогенаогоудетей,что |
|
|
|
||
усиливаетлиполизадипоцитах,накоплениежирныхкислследовательновкр, ви,кет |
|
|
о- |
|||
генез. |
|
|
|
|
|
|
|
Синтез ацетоацетатапроисходиттольков |
митохондриях печени,дацетоацетатл |
||||
либово |
сстанавливаетсядо3 |
-гидроксибутирата,либосп декарбнтннодоацксилируется |
|
е- |
||
тона.Далсоединениявстрие поступаютвкровьразносятсятканям.Ацетон,каклет |
|
|
у- |
|||
чеев ще,удаляетсявыдыхаемымтвовоздухомипотом.Всекетонтеламогутвыдвые |
|
|
е- |
|||
лятьсямочой. |
|
|
|
|
||
|
Используются кетоновыетелавсемитканя,кропечмиэритроцитовени.Особенно |
|
|
|||
активно,даженорме,онипотребляютсямиокардомкоркслнадпочечниковвымем. |
|
|
|
|||
|
Реакцииутилпроисхвзацииобратном.Вцитозоледядке3 |
|
|
-гидроксибутират |
||
окисляется,образующийсяцетоацекаетпронвми,активируетсяохондриизасчетсу |
|
|
к- |
|||
цинил-SКоА ипревацетилращается |
-SКоА,которыйсгораетвЦТК. |
|
|
|||
|
При голодании синкетоновыхтелезускоряется60раз( |
повышение до0,6г/л |
||||
|
принормеменее0,01г/л |
),при |
декомпенсации сахарного диабета I типа – в400 |
|||
|
раздо(г4/л). |
|
|
|
|
|
|
|
|
|
|
|
|
Строениобменлипиедов |
252 |
|
|
З А П А С А Н И Е Ж И Р О В
Реакцбиослипидовнтеза |
могутидти |
клетках всехорганов.Субдлястратоминтеза |
|
||
жиров de novo являетсяглюкоза. |
|
|
|
|
|
Какизвестно,попадаяклетку,глюкозаокисляетсяпо |
|
|
гликолитическомупутидо |
и- |
|
ровинограднкислоты.Пируватмитохондрияхдекарбоксйвацетилируется |
|
|
-SКоА и |
||
вступаетЦТК.Однакосостояниипокоя,приотдыхе,приналичиидостаточногоколич |
|
|
|
е- |
|
стваэнергииклеткереакции |
ЦТК (вчастности,изоцитратдегидр |
огенреакция) зная |
бло- |
||
кируются избытком АТФ и НАДН.Врезультатенакапливпервыйм тЦТКаболитется |
|
– |
|||
цит.Пограконцентрациидиентуперемещаетсявцитозоль,расщепляетсяобразов |
|
|
|
а- |
|
ниемацетил |
-SКоА,которыйдалееиспользуетвбиосинтезехоле теролая |
|
,жирныхкислот |
|
|
триацилглицеролов.

biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
253 |
|
|
|
|
|
|
|
Б И О ЖИСКНРИТНСЕЫЛЗХ |
О Т |
|
Биосижиркислотнтезаиболееыхактивнопроисхцитклетокзоледит |
печени, |
||
кишечника, жировойткани |
всостояниипокилиеды.сле |
|
||
|
Услвыделитьможноэтапабиосинтеза4 : |
|
||
1. Образованиеац тил |
-SКоА изглюкозыиликетогеаминокислот. ных |
|
||
2. Переносацетил |
-SКоА измитохондрийвцитозоль: |
|
||
o |
можетбытьвкомплексекарнитином,такжекакпереносятсявысшжирныекисл |
о- |
||
|
ты,ноздесьтранспортивдругометнаправлении |
, |
||
o |
обычновсоставе |
|
лимоннойкислоты,образующейсявпервойреакцииЦТК. |
АТФ-цитрат-лиазой |
|
Поступающийизмитохондрийцитратвцитозолерасщепляется |
|||
доацетил -SКоА иоксалоацетата . |
|
|||
Оксалоацетат вдальнейшемвосстанавливаетсядомалата,последнийлибоперех |
о- |
|
дитвмитохондриималат( |
-аспартатныйче),либонокдеарбоксилируетсявпируват |
малик- |
ферментом (яблочныйфермент). |
|
|
3. Образование малонил-SКоА изацетил -SКоА. |
|
|
Карбоксилированиеацетил |
-SКоА катализируется ацетил-SКоА-карбоксилазой. |
|

Строениобменлипиедов |
254 |
|
|
4. Синтезпальмитиновойкислоты |
. |
синтазажирныхкислот |
|
|
||
Осуществляетсямультиферментнымкомплексом" |
|
"синоним( |
||||
пальмитатсинтаза)всоставкотороговходитферментов6 ацил |
|
-переносящийбелок |
|
|||
(АПБ). Ацил-переносящпантотеновойбелоквключаетпроизводноекислоты |
|
|
– |
|||
6-фосфопантетеин( |
ФП),имеющий |
HS-группу,подобно |
HS-КоА.Один |
з ферментовко |
м- |
|
плекса, 3 |
-кетоацилсинтаза,такжеимеет |
HS-группу.Взаимодействиеэтихгруппобусловл |
|
и- |
||
ваетначало |
ипродолжени е биосжкислотырнойнтеза,именно |
пальмитиновой кислоты. |
||||
ДляреакцийсинеобходимтезаНАДФН.
Впервых |
двух реакциях (смстр.2 |
77) последовательноприсоедималониляются |
|
-SКоА |
|||
кфосфопантетеинуацил |
|
-переносящегобелка |
(всегда) иацетил |
-SКоА |
кцистеину |
||
3-кетоацилсинтазы (тольков |
первой реакции) |
. 3-Кетоацилсинтазакатализирует |
третью ре- |
||||
акцию – переносацетильнойгруппынаС |
|
2 малониласотщеплениемкарбоксильнойгруппы. |
,дегиопятьдратации |
||||
Далеекетогруппавреакцияхвосстановления |
|
|
приучастииНАДФН |
||||
восстанопревмлращаетилнсобразованиемя тсяовуюасыщенногоацила.Ац |
|
|
|
и- |
|||
лтрансфераза переноегонацис3теинит |
|
-кетоацилсинтазы,кфосфопантетеинуприсоедин |
я- |
||||
етсямалонил |
-SКоА |
ициклповтдообразованряеосптсяльмитиновойтка.слотыя |
|
|
|
||
Пальмитиноваякислотаотщепляетсяшестымферментомкомплексатиоэстеразой. |
|
|
|
||||
|
|
УД Л ЦЕПИНЖ ИРНЫЕ Х |
КИСЛОТ |
|
|
||
Синтезированпальмитикислотапрнеобходимоспоступаетоваяэндоплазми |
|
|
|
а- |
|||
тичрескийтикулум.Здесьучастиеммалонил |
|
|
|
-SКоА иНАДФНцепьудлиняетсядоС |
18 или |
||
С20. |
|
|
|
|
|
|
о- |
Удлимогуиненаятьжирныесыщеннлинолеваяякисо(леот, иноваяы,линолен |
|
|
|
||||
вая)собразпроизвваниемэйккислозандных( отывой |
|
|
|
С19Н39COOH)Но.двойнаясвязь |
|||
животнымиклеткамивводится |
|
|
недалееатома9углерода |
,поэтому |
ω-3и |
||
ω-6-полиненасыщенные жирныекисинлотезируютсяыолькоизсоответствующихпре |
|
д- |
|||||
шественников. |
|
|
|
|
|
|
|
Нап, рахимерк дслотаножетбразоватьсяклеткеаятолькоприна ичии |
|
|
|
и- |
|||
ноленовойилинолевойкислот.Приэтомлинолеваякислота(18:2)дегидрируетсядо |
|
|
|
|
|||
γ-линоленовой(18:3 |
)иудлиняетсядоэйкозотриеновойкислоты(20:3),последняядалее |
|
|
|
|||
вновьдегиддоарахируетсяк доновойслоты(20:4)Такформж. иркислотыныеуются
ω-6ряда .

biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
255 |
|
|
|
|
Дляобразованияжирныхкислот |
|
ω-3-ряда,например,тимнод(20:5),неоновойбход |
и- |
||
моналичие α-линоленовойкисл(18:3),котораятыдегидрируется(18:4),удлиняется(20:4) |
|
||||
опятьдегидрируется(20:5). |
|
|
|
|
|
|
|
Р Е Г У Л Я Ц И Я О Б М Е Н А Ж ИКРИНСЫЛ ХО Т |
|
||
|
|
|
Гормональнаярегуляция |
ин- |
|
Сиокисленнтезтриацжирныхлглицеролкислотзависитсоотновшения |
|
||||
сулин/глюкагон. |
|
|
|
|
|
1. Изменениеколичестваферментов. |
пальмитатсинтазы и ацетил-SКоА-карбоксилаза |
|
|||
Ферментыкомплекса |
являются |
||||
адаптивными ферментами,количествоихвозрастапитанииприусилеуменьшаеом |
|
т- |
|||
сяприголодании |
ипотреблениижира.Индукторомбиосэтферинтезахявляетсяентов |
ин- |
|||
сулин. И,напротив,количествофермента |
|
ТАГ-липазы увеличиваетсяглюкокортикоидами. |
|
||
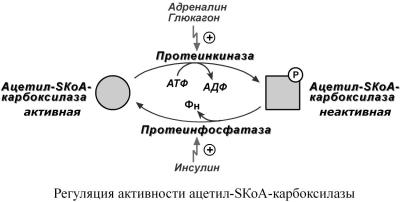
2Ковалентная. модификация. |
|
|
|
||
Благодаря инсулину,глюкагону,адреналину,тиреот енокортикотроопному |
пному |
||||
гормонам происходит ковалентная модификация ферментов ацетил-SКоА-карбоксилазы |
|||||
иТАГ -липазы путемфосфорилирования |
-дефосфорилирования. |
|
|||
Инсулин активируетпротеинфспосдефосфорилированиюбствуетсфатазуи |
акти- |
||||
вации ацетил-SКоА-карбоксилазы.Одновр |
еменновклеткедефосфорилируетсяинактив |
и- |
|||
руетсяТАГ |
-липаза. |
|
|
|
|
Глюкагон, адреналин илидругиегормоны,действуяпоаденилатциклазномумех |
а- |
||||
низму сучастиемцАМФ |
|
-зависимойпротеинкиназы,вызываютфосфорилированиеинг |
и- |
||
бированиеацетил |
|
-SКоА-карбоксилазы, |
лед,овательностанавливаютлипогенез.Одн |
о- |
|
временноониактивируютТАГ |
|
-липазу. |
|
|
|
Строениобменлипиедов |
256 |
|
|
|
При уменьшении количестваинсулинавозрглстании |
юкагона усиливаются липо- |
|||
лиз вжировойткани,поступлениежирныхкислотвпеченьдругиетканиреакцииих |
|
|
|
||
β-окисления. Такоесостояннаблюдаетсяпригипогликемиилюбпр гоисхождения. |
|
|
|||
|
Приобратнсоотношег рмоновначинаюреакцсииж.итезасяиров |
|
|
|
|
|
Врегуляциисинтезаокислеж кислригныхияротриаютучасткаль: |
Метаболическаярегуляция |
|
||
|
|
|
|
||
1Активность. |
ацетил-SКоА-карбоксилазы регулируется: |
|
|||
o |
цитратом – аллостерическийактиваторфермента |
,накапливаетсяцитозолепри |
з- |
||
|
бытколичествеомэн,ергии |
|
|
|
|
o |
ацил-SКоА попринципуобратнойотрицательнойсвязиингибируетфермен, |
|
пре- |
||
|
пятствуясинтезумалонил |
-SКоА.Т.е.если |
ацил-SКоА неуспевступитьэтерифа |
и- |
|
кациюилиусилиполизвклваувеличиваетсяиликепоступлениежирныхки |
|
|
|
с- |
|
лотизвне, автоматблокируетсяновыхнтезчесжирных.слот |
|
|
|
|
|
2. Транспортцитра а |
измитохондриивцитозоль |
подавляется избытком ацил-SКоА,это |
|||
снисинтезжает |
ирныхкислот. |
|
|
|
|
3. Карнитин-ацилтрансфераза ингибируется приобразовании |
|
малонил-SКоА,чтоост |
а- |
||
навливаетпоступациление |
-SКоА внутрьмитохондрий |
снижает β-окисление. |
|
||
Такимобразом, |
усиление синтезажирныхкислотдостигаетсяприпоступлении |
|
цитра- |
||
та и при наличии малонил-SКоА.Послесинтезамолекулыацил |
|
-SКоА быстропоступаютна |
|
||
этерификациюглицероладоТАГненак вцитозолепливаются. |
|
|
|
|
|
Если вклеткенедостаточноэнергии |
,тонеобходактивирмовать |
|
β-окислениежи |
р- |
|
ныхкислслучает.Вэтогормонавлиянвызываютлиьныеилиполизя(поступление |
|
|
|
|
|
жирныхкислотизвне)накоплениеацил |
|
-SКоА вцитозоле,чтоавтоматически,через |
|
||
уменьшениеколичествацитратамалонил |
|
-SКоА (см пункт2), |
препятствсинтезует |
о- |
|
выхжирныхкислот.Параллельносм(пункт3) |
|
убымальонил |
-SКоА иактивация карнитин- |
||
ацил-трансферазызапускает |
β-окисление. |
|
|
|
|
Попавшаямитохожиркислндриюеизбежноаятакисляетсядоацетил |
|
|
-SКоА. |
|
|
При появлении избытка ацетильныхгрупп |
инакопленииАТФ |
|
в митохондриях печени |
||
ониперенасинтезправляются |
кетоновыхтел. |
|
|
|
|

biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
257 |
|
|
|
|
С И НТТРЕИЗА Ц И Л Г Л И Ц Е Р О |
Л О В |
|
|
|||
ИФ О С Ф О Л И П И Д О В |
|
|
|
|||
ОБ ЩПРИНЦИПЫБИОСИНЕ |
|
ТЕЗА |
|
|
|
|
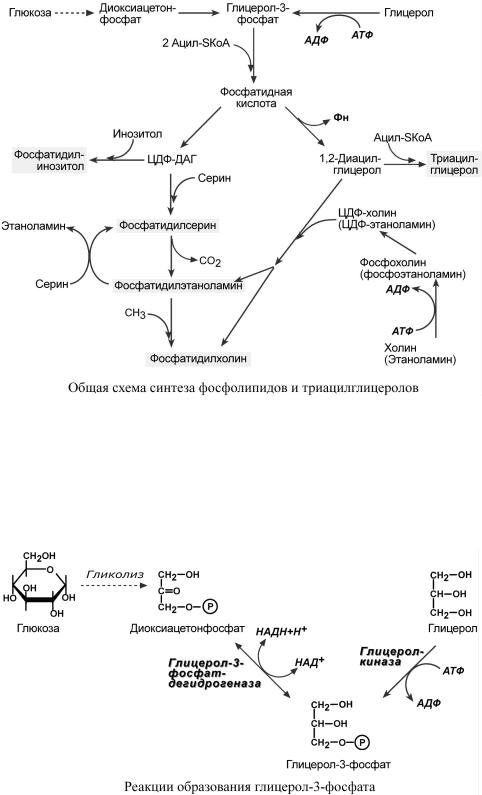
Начальреакцииситриацилглицероловнтезаыефосфолипсов роисхадаютдов |
|
|
|
|
|
о- |
дятпр иналичии глицерола и жирныхкислот |
. |
|
|
|
|
|
Вреакцияхбиосинтможновыдследующиезалить |
|
|
события: |
|
|
|
1. Образование глицерол-3-фосфата изглюкозы |
черездиоксиацетонфосфат |
или при фос- |
||||
форилировании свободногоглицер |
ола. |
|
|
|
|
|
2.. Биосинтезфосфатиднойкислоты |
– требует наличияглицерол |
-3-фосфата ижирныхки |
с- |
|||
лот. Присвязыванииглицерол |
-3-фосфата сжирнымикисинтезлотамируется |
|
фосфа- |
|||
тиднаякислота |
. Далеефосфатиднаякислота |
можпредвумявращатьсяпут ми |
|
– в |
||||
ЦДФ-ДАГ дефосфорилироватьсядо |
1,2-ДАГ. |
|
|
|||||
3. Синтезтриаци |
|
лглицерола – идетиз1,2 |
-ДАГ последефосфорилирования |
фосфатидной |
||||
кислоты. |
Образованный 1,2-ДАГ ацилируетсядоТАГ. |
|
|
|
||||
4. Синтезфосфолипидов |
|
. |
|
|
|
|
||
Сейчасрассматриваютсядва |
пути синтезафосфолипидов. |
|
|
|||||
Поодномупути |
|
1,2-ДАГ непревраща |
етсявТАГ,а |
связывается сэтаноламином |
с обра- |
|||
зованием фосфатидилэтаноламина, либо схолином |
– образуется фосфатидилхолин. По |
|||||||
другомупути |
, ЦДФ-ДАГсвязываетсялибоиноз,лсериномбообразованиемлом |
|
|
о- |
||||
ответственно фосфатидилинозитола или фосфатидилсерина. |
|
|
||||||
Придекарбоксил |
ированиифосфатидилсерина |
далее образуется фосфатидилэтанола- |
||||||
мин,который |
может пре,всвоюратитьсяочередь, |
|
фосфатидилхолин. |
|
|
|||
Синтезированныйлюбымспособом |
|
фосфатидилэтаноламин также способен взаимо- |
||||||
действовать ссерином |
|
обратно образовывать фосфатидилсерин. |
|
|
||||
Такимобр, изждыйзомосновныхфосфолипидов |
|
– фосфатидилсерин, |
|
- |
||||
этан,фоламинсфатидилхолин |
|
– способенпоступатьизразныхисточников,чтоблагоприя |
|
т- |
||||
ствуетподдержаниюребалансауемого. |
|
|
|
|
|
|
||
Строениобменлипиедов |
|
|
258 |
|
|
|
|
ЦДФ-ДАГ, являясьактивнойформойфосфа |
|
тиднойкислоты, |
способен превращаться не |
только в указанные,ноивдругие |
фосфолипиды, |
например, |
в кардиолипин. |
|
О Б Р АГЗЛОИВЦАЕНРИОЕЛ |
- 3 - Ф О С Ф А Т А |
|
|
Вначале всегопроцесобразованиепроисходитглицерол |
-3-фосфата. |
|
||
Глицерол впечени |
фосфорилируется вреакц |
иис спользованиемАТФ.В |
мышцах, |
|
жировойткани |
идругихданная |
реакцияотсутс,поэвнихтглицеролвуетому |
|
-3-фосфат б- |
разуетсяиз |
диоксиацетонфосфата,метаболитагликолиза. |
|
|
|
С И НФТОЕСЗКФИА Т И Д Н О Й |
С Л О Т Ы |
Жирныекислоты |
,поступающие изкровип аспадеХМ,ЛПОНП |
илисинтезир о- |
ванныеклеткеизглюктакжедоактивироватьсялжнызы.Онипре ацилращаются |
-SКоА |
|
вАТФ -зависимойреакции. |
|
|

biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
259 |
|
|
|
|
Приналичииглицерол |
-3-фосфатаиацил |
-SКоА синтезируется фосфатиднаякислота |
. |
|
Взависимоотвидажирнойки,образующаясятилотыфосфатиднаякислотаможет |
|
|
||
содержатьнасыщеннилиненасжирныекислотщенн, ые |
|
внекородеэтором |
опреде- |
||
ляет ее дальнейшуюсудьбу: |
|
|
|
||
o |
еслииспользуютсянасыщенмонокислотыненыеп(асыщенныельмитиновая |
|
,ст е- |
||
|
ариновая, |
пальмитолеиновая,олеиновая),тофосфатиднаякислотанапрсивляется |
|
н- |
|
|
тезТАГ, |
|
|
|
|
o |
привключеполижирнииенакилинолес(ылотхщен,арахидоновая, ыховая |
|
|
||
|
кислотыомега |
-3ряда)фосфатиднаякислотаявляпретсядшефос.фолипидовтвенником |
|
|
|
|
|
С И Н Т Е ЗТ Р И А Ц И Л Г Л И Ц Е Р О Л О В |
|
||
|
БиосинтзаключаетсяТАГвд фосфорфосфатиднк лированиислотыприсоедй |
|
и- |
||
ненацильнойгруппы. |
|
|
|
|
|
|
БиосинТАГклеткамиез |
увеличиваетприсоблюхотябыдизногоенииследующихя |
|
|
|
условий: |
|
|
|
|
|
o |
наличиеисточникадешевой" "энергии |
.Например, 1) |
диетабогатая |
простыми уг- |
|
|
леводами (глюкоза,сахароза) |
– приэтконцентрациямглюквкрпослеедывизыре |
|
з- |
|
|
коповышаетсяиподвлияниемнсулактпровнонасинтезжировсходит |
|
|
адипоци- |
|
|
тах и печени. 2) наличие этанола приуслнормальногвии |
опитания,примеромможет |
|
||
|
служить" |
пивное ожирение", |
|
|
|
o |
повышениеконцентрациижирныхкислот |
вкрови |
– синтез происходвпечен, ит |
||
|
нап, русиленноммерлиполизевжироклеткахподвыхоздействиемкаких |
|
-либо |
||
|
веществфармпр( ,кофит..е),припаратыэмоциин |
ональномстреиотсутствиисе(!) |
|
||
|
мышечнойактивности, |
|
|
|
|
o |
высокиеконцентрацииинсулина |
инизкиеконцентрацииглюкагона |
|
– послеприема |
|
|
пищи. |
|
|
|
|