

Тип Плеченогие (Brachiopoda)
Тело одето однослойным кожным эпителием, под которым находится тонкий слой соединительной ткани, а затем мерцательный перитонеальный эпителий, выстилающий обширную вторичную полость тела. Сплошной кожно-мускульный мешок отсутствует вследствие развития раковины, которая не позволяет телу сокращаться и изменять форму, но сильно развиты отдельные мышечные пучки, идущие через полость тела от внутренней поверхности одной створки к другой (Шарова, 2004).
Целом плеченогих представлен обширной полостью тела, содержащей кишечник и пересекаемой мышцами раковины. Мезентерий делит целом на правую и левую половины; целом дает выросты, проникающие в толщу мантийных складок, а также узкие каналы, проходящие внутри рук. От этих каналов отходят тонкие веточки в каждое щупальце. В полости тела плеченогих, кроме продольных мезентериев, имеются и неполные поперечные перегородки. Иногда их рассматривают как остатки настоящих диссепиментов, разделяющих целом на три отдела: передний - эпистомальный; средний, посылающий целомические каналы в лофофор и щупальца, и задний - туловищный. Полость тела содержит бесцветную жидкость с рассеянными в ней амебоидными клетками (Островерхова, 2005). Подробное внутреннее строение целома плеченогих показано на рис. 4.
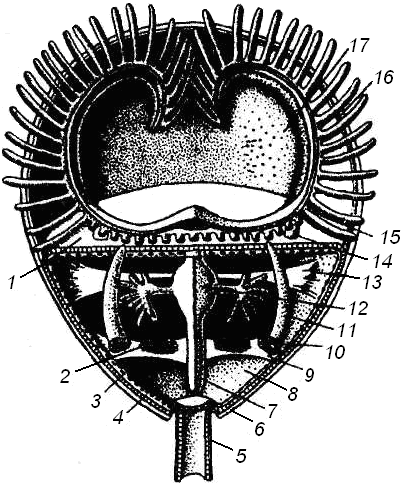
Рис. 4. Схема строения плеченогого (Догель, 1981): 1 - передняя стенка тела; 2 - яичник; 3 - перитонеальный эпителий; 4 - кожный эпителий; 5 - стебелек; 6 - раковина; 7 - средняя кишка; 8 - целом; 9 - задняя складка перитонеального эпителия; 10 - воронка целомодукта; И - целомодукт; 12 - желудок; 13 - передняя складка перитонеального эпителия; 14 - основание руки; 15 - пищевая бороздка между основаниями щупалец; 16- спинная мантийная складка; 17- мантийная полость
Развитие целомической полости у трохофорных животных
Трохофорные, или членистые, животные (Trochozoa) включают типы: Моллюски (Mollusca), Сипункулиды (Sipunculida), Эхиуриды (Echiurida), Кольчатые черви (Annelida), Тихоходки (Tardigrada), Онихофоры (Onychophora), Членистоногие (Arthropoda). Для большинства из них характерна пелагическая личинка - трохофора, обладающая слепым или сквозным кишечником, первичной полостью тела, протонефридиями. В силу своеобразия и преобладающего среди трохозоев числа целомических животных вопрос сместился в сторону выяснения их происхождения.
У трохофорных животных, эволюция которых была направлена по пути развития метамерии, целом в разных группах Trochozoa то приобретал, то терял метамерное строение. В связи с этим в эволюции трохозоев выделяется три основных направления, представленных соответствующими группами, характеризующимися функциональными преобразованиями целома и разной степенью развития метамерии.
Первая группа характеризуется исходным нерасчлененным состоянием целома, не связанным с метамерией тела. Современные Echiurida, Priapulida, вероятно, являются реликтовыми группами таких первично несегментированных Coelomata, причем целом у них возник независимо путем эпителизации стенок схизоцельных (первичных) опорных полостей. Схизоцельный целом у животных появляется раньше, чем развивается истинная метамерия.
Вторая группа представлена в основном аннелидами (Annelida), имеющими разную степень и формы расчленения целома. В ряду трохофорных животных развитие метамерного целома кольчецов больше всего связано с развитием метамерии тела, физиологией, экологией и особенностями способов движения. У полихет, олигохет и пиявок можно наблюдать несколько разных способов движения и соответственно разное строение целома.
Так, развитие многочисленных парных целомических камер у свободноживущих полихет функционально связано с изгибанием их тела в горизонтальной плоскости и сопровождается развитием параподиальных выростов. Преобладание перистальтического способа движения у роющих полихет и олигохет сопряжено с частичной утратой поперечных перегородок полости тела и в ряде случаев параподиальных выростов. У пиявок преобладает изгибание тела в вертикальной плоскости. При таком способе движения парные целомические камеры теряют свое функциональное значение. Опорная функция в этом случае переходит к паренхиме, а гомеостатическая, транспортная и выделительная функции сохраняются за целомической полостью, которая приобретает форму лакунарной системы.
Появление целома у предков современных целомических животных - это ароморфоз, открывший новые пути для адаптивной радиации многообразной группы Coelomata. По поводу источников происхождения целома существует несколько теорий: схизоцельная, миоцельная, гоноцельная и энтероцельная. Основные различия между этими теориями (гипотезами) заключаются в том, какая из многих функций целома принимается за первичную и какой процесс формирования целома принимается за первичный (Островерхова, 2005).
Схизоцельная теория рассматривает целом как гомологичное образование первичной полости тела Nemathelminthes, представляющее простое ее усовершенствование. Эта теория не объясняет, почему при эмбриональном развитии у многих животных целом формируется за счет стенок кишечника.
По миоцелъной теории констатируется образование целома как полости в мускульном зачатке животного, которая заполнилась жидкостью и выполняла опорную функцию для окружающей мускулатуры. Другие функции целома эта теория не объясняет (Догель, 1981).
По гоноцельной теории целом производится из половых желез низших червей. Согласно этой теории, полость каждого целомического мешка соответствует полости гонады турбеллярии или не-мертины, перитонеальный эпителий происходит из стенки половой железы, а целомодукты - из половых протоков. В этой теории много натяжек.
Сторонники энтероцелъной теории считают, что целом берет начало от гастроваскулярной системы кишечнополостных (Coelenterata) и гребневиков (Ctenophora), которые рассматриваются как отдаленные предки целомических животных. Так как у высших кишечнополостных под энтодермой гастроваскулярных каналов развиваются гонады, то энтероцельная теория хорошо объясняет половую функцию целома. Она, по существу, включает в себя и гоноцельную теорию (Островерхова, 2005).