

43.Особенности регуляции в кишечной стенке и других тканях биосинтеза хс; роль гормонов: инсулина, т3,т4, витамина рр.
Регуляция синтеза ХС
Ключевой фермент синтеза ХС ГМГ-КоА-редуктаза регулируется несколькими способами:
В печени синтез редуктазы подвержен суточным колебаниям: максимум на полночь и минимум на утренние часы.
ХС, желчные кислоты (в печени) репрессируют ген ГМГ-КоА-редуктазы. В норме поступление ХС с пищей снижает синтез собственного ХС в печени, однако с возрастом эффективность этой регуляции у многих людей снижается и уровень ХС повышается.
Инсулин через дефосфорилирование осуществляет активацию ГМГ-КоА-редуктазы.
Глюкагон через фосфорилирование осуществляет ингибирование ГМГ-КоА-редуктазы.
Повышение концентрации исходного субстрата ацетил-КоА стимулирует синтез ХС.
Таким образом, синтез ХС активируется при питании углеводами и ингибируется при голодании.
Основная роль витамина РР в организме – участие в окислительно-восстановительных процессах. Витамин РР способствует нормальному росту тканей, оказывает благотворное влияние на жировой обмен, участвует в преобразовании сахара и жиров в энергию, снижает в крови уровень «плохого» холестерина.
Активность ГМГ-редуктазы возрастает при введении инсулина и тиреоидных гормонов(Т3 и Т4), это приводит к усилению синтеза ХС.
При голодании, тиреоидэктомии, введении глюкагона и глюкокортикоидов – угнетение синтеза ХС.
44.Реакции образования и распада эфиров холестерина – роль ахат и гидролазы эхс, особенности тканевого распределения хс и его эфиров, значение.
Этерификация ХС
ХС образует с ЖК сложные эфиры (ЭХС), которые более гидрофобны чем сам ХС. В клетках эту реакцию катализирует АХАТ (ацилКоА: холестеролацилтрансферазой):
ХС + АцилКоА → ЭХС + HSKoA
АХАТ содержится лишь в некоторых тканях, синтезированный им ЭХС формирует в цитоплазме липидные капли, которые являются формой хранения ХС.
По мере необходимости ЭХС гидролизуются холестеролэстеразой на ХС и ЖК.
ЭХС синтезируются также в крови в ЛПВП под действием ЛХАТ (лецетин: холестеролацилтрансферазой):
ХС + лецитин → ЭХС + лизолецитин
В печени синтезируется более 50% ХС, в тонком кишечнике — 15— 20%, остальной ХС синтезируется в коже, коре надпочечников, половых железах. С пищей поступает в сутки 0,3—0,5г (0.3-0.4) ХС. Общее содержание ХС в организме составляет в среднем 140г, 90-93% находиться в клетках, 7-10% - в крови (5,2+1,3 ммоль/л).
В составе ЛП ЭХС обеспечивают большую часть транспорта ХС в крови. На долю ЭХС крови приходиться 75% от общего количества ЭХС в организме.
Эфиры холестерина являются основной формой запасов холестеринов и должны подвергнуться гидролизу до свободного холестерина, который используется в стероидогенезе.
45.Катаболизм хс, тканевые особенности, пути удаления из организма. Лекарственные препараты и пищевые вещества, снижающие содержание хс в крови.
Выведение ХС из организма
Клетки организма, за исключением печени, не перерабатывают холестерин. Небольшое количество холестерина используется для выработки гормонов в надпочечниках, яичниках и мошонке, часть его превращается в желчные кислоты.
Так как производные циклопентанпергидрофенантрена (стероиды) водонерастворимы и в организме не расщепляются, они выводятся из организма в основном с калом в составе желчи и немного с потом через кожу.
В сутки из организма выводится от 1,0г до 1,3г ХС. ХС выводится с желчью (0,5-0,7 г/сут) в основном в виде жёлчных кислот и частично в чистом виде. Часть ХС в кишечнике под действием ферментов бактерий восстанавливается по двойной связи, образуя холестанол и копростанол. С кожным салом в сутки выделяется 0,1г ХС.
Витамин В3 (ниацин) — расширяет сосуды, обладает антиатерогенным действием, так как понижает уровень холестерола и триглицеридов.
Оливковое масло и некоторые другие продукты, такие, как орехи, авокадо, масло канола и арахисовое масло, имеют высокое содержание абсолютно другого жира — мононенасыщенного.
бобы и другие бобовые содержат растворимую в воде клетчатку под названием пектин, который окружает холестерин и выводит его из организма
Фрукты, морковь, чеснок (но блин если его много есть,то лишитесь друзей) также обладают способностью снижать холестерин из-за наличия в них пектина.
Ниацин, Витамин С, Витамин Е, Кальций способствуют выведению ХС.
Лекарственные средства:
Вазилип,Зокор,Липримар,Крестор,Симгал.
Реакции биосинтеза кетоновых тел, регуляция, значение.
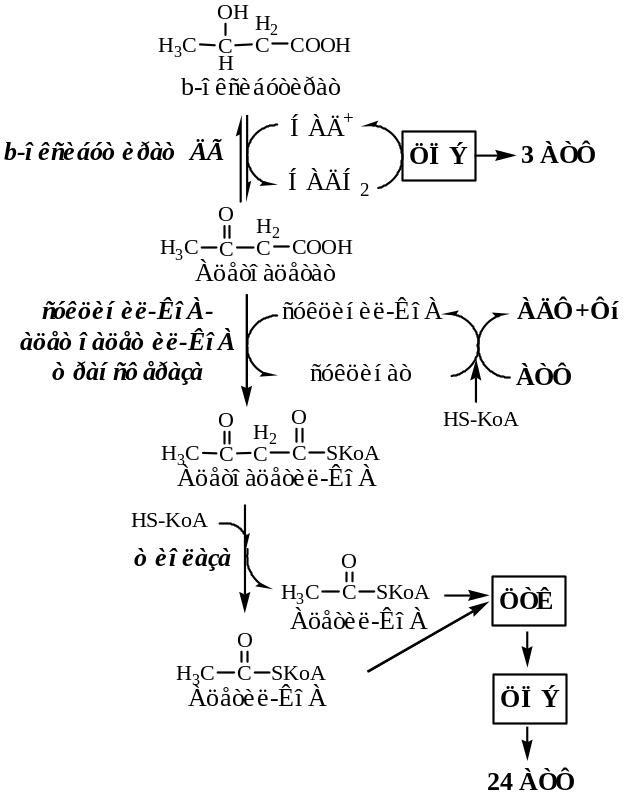
К кетоновым телам (КТ) относят β-оксибутират, ацетоацетат и ацетон.
Синтез КТ
β-оксибутират и ацетоацетат синтезируются в митохондриях печени из ЖК. Ацетон образуется в крови неферментативно:
Реакции:
Под действием тиолазы 2 ацетил-КоА взаимодействуют с образованием ацетоацетил-КоА;
Под действием ГМГ-КоА-синтазы с ацетоацетил-КоА взаимодействует третья молекула ацетил-КоА, образуя 3-гидрокси-3-метилглутарил-КоА (ГМГ-КоА);
ГМГ-КоА-лиаза катализирует расщепление ГМГ-КоА на свободный ацетоацетат и ацетил-КоА;
Высокая концентрация НАДH2, образованная при активном β-окислении ЖК, восстанавливает в печени большую часть Ацетоацетата до β-оксибутирата. Фермент β-гидроксибутират ДГ;
Ацетоацетат и β-гидроксибутират выделяются в кровь;
При высокой концентрации в крови ацетоацетата часть его неферментативно декарбоксилируется, превращаясь в ацетон.
Регуляция синтеза КТ
Глюкагон в жировой ткани активируется распад ТГ. ЖК поступают в печень в большем количестве, чем в норме, что увеличивает скорость их β-окисления.
Глюкагон в печени направляет ЩУК на глюконеогенез, подавляя ЦТК. Образующийся из ЖК ацетил-КоА не окисляться в ЦТК, накапливается в митохондриях и идет на синтез КТ.
Регуляторный фермент синтеза КТ — ГМГ-КоА синтаза. Синтез ГМГ-КоА синтазы индуцируют высокие концентрации ЖК, ингибируют высокие концентрации НSКоА. Избыток ЖК в печени связывает НSКоА, концентрация НSКоА снижается, ГМГ-КоА-синтаза активируется. И наоборот, дефицит ЖК в печени увеличивает концентрацию НSКоА, фермент ингибируется.
Биологическая роль КТ
КТ — хорошие топливные молекулы, окисление β-гидроксибутирата до СО2 и Н2О обеспечивает быстрый синтез 26 молекул АТФ. Окисление КТ, как и ЖК сберегает глюкозу, что имеет большое значение в энергоснабжении аэробных тканей при длительном голодании и физических нагрузках, когда возникает дефицит глюкозы. Для нервной ткани КТ имеют исключительное значение, так как в отличие от мышц и почек, нервная ткань практически не использует ЖК в качестве источника энергии(ЖК не проходят гематоэнцефалический барьер).
Преимущество КТ перед ЖК: 1) КТ водорастворимы, а ЖК – нет; 2) ЖК разобщают окислительное фосфорилирование и усиливают синтез ТГ, а КТ – нет.
Ацетон, в отличие от β-оксибутирата и ацетоацетата, не утилизируется тканями. Он выделяется с выдыхаемым воздухом, мочой и потом, что позволяет организму избавляться от избытка КТ, которые не успели вовремя окисляться.
Кетоацидоз
В норме концентрация КТ в крови составляет 1—3 мг/дл (до 0,2 мМоль/л), но при голодании значительно увеличивается. Увеличение концентрации КТ в крови называют кетонемией. При кетонемии развивается кетонурия - выделение КТ с мочой. Накопление КТ в организме приводит к кетоацидозу, так как КТ (кроме ацетона) являются водорастворимыми органическими кислотами (рК~3,5).
Ацидоз достигает опасных величин при сахарном диабете, так как концентрация КТ при этом заболевании может доходить до 400—500 мг/дл. Тяжёлая форма ацидоза — одна из основных причин смерти при сахарном диабете.
Реакции распада кетоновых тел до Ацет-КоА и, далее до СО2 и Н2О, схема, энергетический баланс.
Окисление КТ в периферических тканях
Как и ЖК, КТ окисляются только в аэробных условиях, обеспечивая синтез АТФ.
β-Гидроксибутират, попадая в клетки, дегидрируется НАД-зависимой дегидрогеназой и превращается в ацетоацетат. НАДН2 направляется ЦПЭ;
Сукцинил-КоА-ацетоацетат-КоА-трансфераза активирует ацетоацетат, при переносе КоА с сукцинил-КоА на ацетоацетат. Этот фермент не синтезируется в печени, поэтому печень не использует КТ как источники энергии;
Тиолаза расщепляет ацетоацетил-КоА на 2 Ацетил-КоА, которые направляются в ЦТК.
КТ — хорошие топливные молекулы, окисление β-гидроксибутирата до СО2 и Н2О обеспечивает быстрый синтез 26 молекул АТФ. Окисление КТ, как и ЖК сберегает глюкозу, что имеет большое значение в энергоснабжении аэробных тканей при длительном голодании и физических нагрузках, когда возникает дефицит глюкозы. Для нервной ткани КТ имеют исключительное значение, так как в отличие от мышц и почек, нервная ткань практически не использует ЖК в качестве источника энергии(ЖК не проходят гематоэнцефалический барьер).
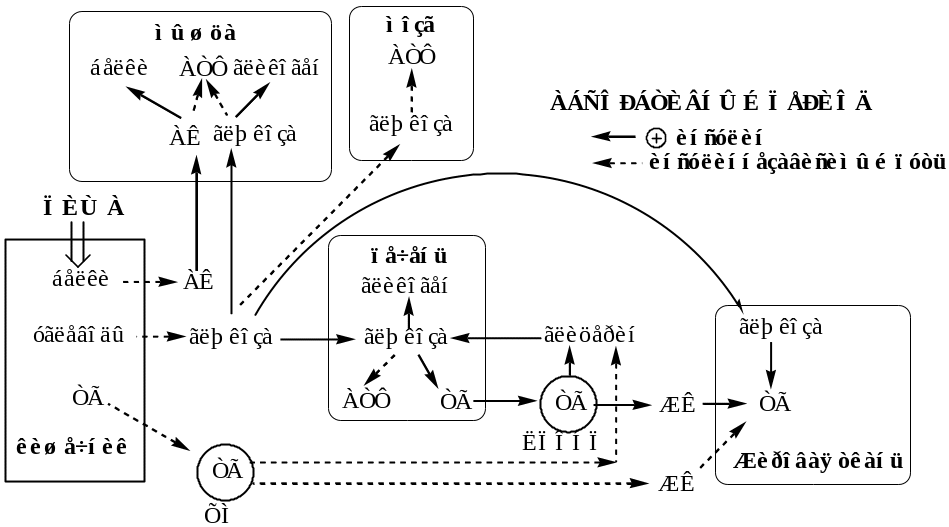
Интеграция липидного и углеводного обменов – роль печени, жировой ткани, кишечной стенки и др.
Межорганный уровень регуляции липидного обмена (Глюкозожирнокислотный цикл - цикл Рендла).
Абсорбтивный период
Абсорбтивным называют период пищеварения. В абсорбтивный период происходит поступление с пищей глюкозы, аминокислот и ТГ. Процесс пищеварения и высокая концентрации в крови глюкозы и аминокислот активирует секрецию инсулина и снижает секрецию глюкагона в поджелудочной железе.
Инсулин стимулирует использования метаболитов для запасания энергоносителей. Он активирует синтез гликогена в мышцах и печени, ТГ - в печени и жировой ткани, белков – в мышцах и печени.
Инсулин подавляет глюконеогенез в печени, липолиз в жировой ткани, катаболизм белков в мышцах и печени.
Главный потребитель глюкозы - печень, она фиксирует до 60% всей поступившей глюкозы. Синтезированные в печени и жировой ткани ТГ запасаются в основном в жировой ткани. Биосинтез ЖК de novo в жировой ткани человека протекает с высокой скоростью только после предшествующего голодания.
 Постабсорбтивным
называют период после завершения
пищеварения до следующего приёма пищи.
Голодание - это состояние, когда
пища не принимается в течение суток и
более.
Постабсорбтивным
называют период после завершения
пищеварения до следующего приёма пищи.
Голодание - это состояние, когда
пища не принимается в течение суток и
более.
Постабсорбтивный период
Снижение в постабсорбтивный период концентрации в крови глюкозы подавляет секрецию инсулина и стимулирует секрецию глюкагона в поджелудочной железе.
Глюкагон ускоряет процессы мобилизации депонированных энергоносителей. Он стимулирует липолиз в жировой ткани, гликогенолиз (гликоген расходуется за 18—24 ч), глюконеогенез (активируется через 4—6 ч после приема пищи), β-окисление ЖК в печени, катаболизм белков в мышцах и печени, запускает в печени синтез КТ из ЖК.
Глюкагон подавляет синтез гликогена в мышцах и печени, ЖК и ТГ - в печени и жировой ткани, белков – в мышцах и печени.
Метаболизм направлен, главным образом, на поддержание концентрации в крови глюкозы, которая служит основным энергетическим субстратом для мозга и единственным для эритроцитов.
В результате в крови повышается количество ЖК (в 2 раза), которые становятся важными источниками энергии для печени, мышц, сердца, почек, и жировой ткани. Т1/2 ЖК в крови тоже очень мал (менее 5 мин), что означает существование быстрого потока ЖК из жировой ткани к другим органам.
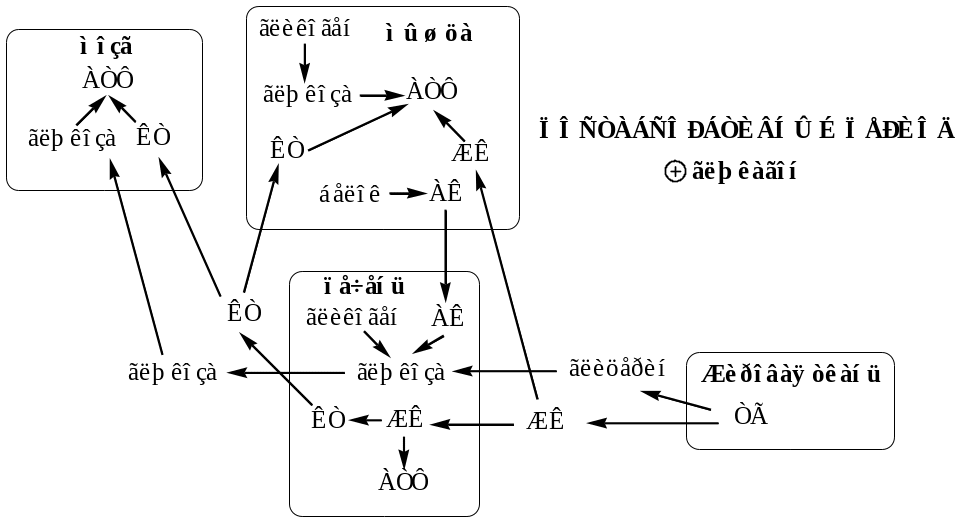
Уровни и механизмы регуляции обмена липидов (перечислить).