

36. Пальмитилсинтетазный комплекс: структура, субклеточная локализация, функция, регуляция, последовательность реакций одного оборота процесса, энергетический баланс.
Синтез ЖК происходит в абсорбтивный период.
Гликолиз + окислительное декарбоксилирование пирувата => АсКоА увелич. в МТХ
А синтез то ЖК в цитозоле! => осуществляется перенос АсКоА через внутреннюю мембрану МТХ
цитратсинтаза
АсКоА + Оксалоацетат Цитрат + HS-KoA
Транслоказа переносит в цитоплазму:
ПВК идет обратно в МТХ
НАДФН – источник Н+ (+ еще идет из ПФП)
АсКоА – исходный субстрат для синтеза ЖК
Образуется малонил-КоА (регуляторная реакция)
Ацетил-КоА-карбоксилаза
АсКоА + СО2 + АТФ НООС-СН2-СО~S-КоА + АДФ + Н3РО4
малонил
Дальше синтез продолжается на мультиферментативном комплексе – пальмитоилсинтетазе. Он состоит из 7 центров, обладающих разными каталитическими активностями. Комплекс последовательно удлиняет радикал ЖК на 2 атома углерода.
Последовательность реакций: (БЛЕЯТЬ! Ну и херь дальше!!!)
Перенос ацетила с КоА на SH-группу цистеина ацетилтрансацилазным центром;
Перенос малонила с КоА на SH-группу АПБ малонилтрансацилазным центром;
Кетоацилсинтазным центром ацетильная группа конденсируется с малонильной с образованием кетоацила и выделением СО2.
Кетоацил восстанавливается кетоацил-редуктазой до оксиацила;
Оксиацил дегидратируется гидратазой в еноил;
Еноил восстанавливается еноилредуктазой до ацила.
В результате первого цикла реакций образуется ацил с 4 атомами С (бутирил). Далее бутирил переносится из позиции 2 в позицию 1 (где находился ацетил в начале первого цикла реакций). Затем бутирил подвергается тем же превращениям и удлиняется на 2 атома С (от малонил-КоА).
Аналогичные циклы реакций повторяются до тех пор, пока не образуется радикал пальмитиновой кислоты, который под действием тиоэстеразного центра гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту.
Суммарное уравнение синтеза пальмитиновой кислоты из ацетил-КоА и малонил-КоА имеет следующий вид:
CH3-CO-SKoA + 7 HOOC-CH2-CO-SKoA + 14 НАДФН2 → C15H31COOH + 7 СО2 + 6
Н2О + 8 HSKoA + 14 НАДФ+
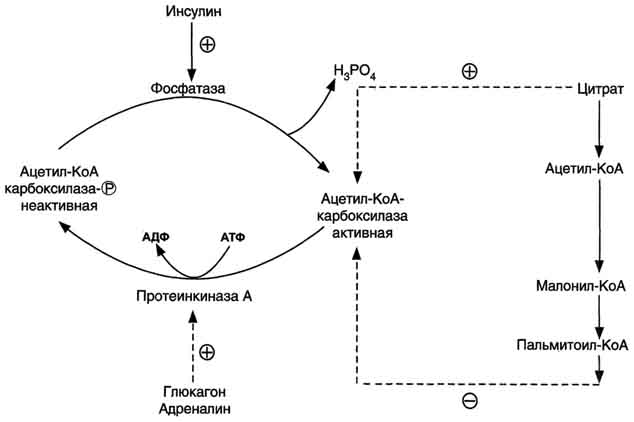
Регуляция:
Самого регуляторного фермента – ацетил-КоА-карбоксилазы
Диссоциация комплексных субъединиц фермента – неакт, а ассоциация – акт
Активатор: цитрат
Ингибитор: пальмитоил-КоА
А) Фосфорилирование фермента – неакт
Это в постабсорбтивный период, при физической работе → глюкагон, адреналин →аденилатциклазная система → активируется протеинкиназа А → фосфорилирование
Б) Дефосфорилирование фермента – акт
Абсорбтивный период → инсулин → активирует фосфатазу → под действием цитрата поляризация фермента → он активируется
37. Реакции удлинения – укорочения жирных кислот, субклеточная локализация ферментов.
Элонгация - удлинение ЖК.
ЖК могут синтезироваться в результате удлинение пальмитиновой кислоты и других более длинных ЖК.
для каждой длины ЖК существуют свои элонгазы.
идет в аЭПС
субстрат – любые ЖК
вместо синтетазы ЖК работает в качестве фермента КоА
продукт элонгации в гепатоците – стеариновая кислота, в нейронах – цереброновая, лингоцериновая ЖК
Удлинение цепи жирной кислоты происходит путем последовательного присоединения к соответствующему ацил-КоА двууглеродных фрагментов при участии малонил-КоА и НАДФН
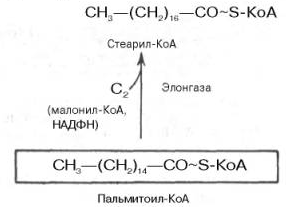
Более подробная схема – учебник Северена (синий), стр. 416