

Сро: стадии и факторы инициации, реакции образования активных форм кислорода.
Кислород в составе молекулы Н2О стабилен, потому что его внешняя электронная орбита укомплектована е- (Н:О:Н). Химические соединения, в составе которых кислород имеет промежуточную степень окисления, имеют высокую реакционную способность и называются активными формами кислорода (АФК). К активным формам кислорода относятся свободные радикалы кислорода и перекиси. Свободный радикал - свободный атом или частица с неспаренным электроном.
Образование активных форм кислорода
АФК во многих клетках образуются в основном в ферментативных и неферментативных реакциях в результате последовательного присоединения е- к кислороду:
О2 + 1е- → О∙2 супероксидный анион-радикал (˙О::О:).
О∙2 +1е- → О2-2 пероксидный анион (:О::О:), он быстро протонируется с образованием перекиси водорода О2-2 + 2Н+ → Н2О2 (Н:О::О:Н)
Н2О2 + 1е- → НО∙ + ОН- гидроксильный радикал, ОН- протонируется с образованием воды ОН- + Н+ → Н2О
ОН∙ + 1е- → Н2О (Н:О:Н)
Ферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать ЦПЭ. Утечка е- из ЦПЭ на кислород является основным путем образования АФК в большинстве клеток:
В цепи окислительного фосфорилирования Q принимая 1 е- превращается в свободный радикал семихинон НQ∙, который при реоксигенации ишемических тканей может непосредственно взаимодействовать с кислородом, образуя супероксидный анион-радикал: HQ· + O2 → Q+ О∙2 + H+;
в монооксигеназных реакциях е- с цитохрома Р450 переходит на кислород с образованием супероксидного анион-радикала, который иногда теряется с активного центра.
Аэробные дегидрогеназы (ФАД-зависимые оксидазы) переносят е- и Н+ с субстрата на кислород с образованием перекиси водорода. Примеры таких оксидаз — оксидазы аминокислот, супероксид дисмутаза, оксидазы, локализованные в пероксисомах.
Неферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать:
1). Металлы переменной валентности. Наличие в клетках Fe2+ или ионов других переходных металлов катализирует образования АФК. Например, в эритроцитах окисление иона железа гемоглобина способствует образованию супероксидного анион-радикала.
Hb(Fe2+) + O2 → MetHb(Fe3+) + О∙2
H2O2 + Fe2+ → Fe3+ + HO- + HO· (реакция Фентона)
HOCl + Fe2+ → Fe3+ + Cl- + HO· (реакция Осипова)
2). Радикалы. АФК, обмениваясь электроном, легко переходят друг в друга: О∙2 + Н2О2 → О2 + НО∙ + ОН-
Реакции образования продуктов ПОЛ, используемых для клинической оценки состояния ПОЛ.(ребята, сами реакции я тупо не нашел, но нашел неплохо изложенные этапы ПОЛ)
Стадии перекисного окисления липидов
1) Инициация: образование свободного радикала (L•)
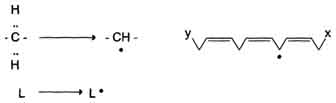
Инициирует реакцию чаще всего гидроксильный радикал, отнимающий водород от СН2-групп полиеновой кислоты, что приводит к образованию липидного радикала.
2) Развитие цепи:
L • + О2 → LOO • LOO• + LH → LOOM + LR•
Развитие цепи происходит при присоединении О2, в результате чего образуется липопе-роксирадикал LOO• или пероксид липида LOOH.
ПОЛ представляет собой свободнорадикальные цепные реакции, т.е. каждый образовавшийся радикал инициирует образование нескольких других.
3) Разрушение структуры липидов
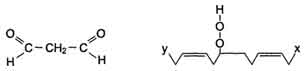
Конечные продукты перекисного окисления полиеновых кислот - малоновый диальдегид и гидропероксид кислоты.
4) Обрыв цепи - взаимодействие радикалов между собой:
LOO• + L• → LOOH + LH L• + vit E → LH + vit E• vit E• + L• → LH + vit Еокисл.
Развитие цепи может останавливаться при взаимодействии свободных радикалов между собой или при взаимодействии с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в стабильную окисленную форму.