

Особенности метаболизма белой жировой ткани
Энергетический обмен низкий, преимущественно анаэробный, ткань потребляет мало кислорода. Энергия АТФ в основном тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина).
Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для собственных нужд. На экспорт в жировой ткани синтезируются лептин, белки острой фазы воспаления (α1-кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С3, фактор В), интерлейкины.
Углеводный обмен. Невысокий, преобладает катаболизм. Углеводный обмен в жировой ткани тесно связан с липидным.
Липидный обмен
Жировая ткань стоит на 2 месте по обмену липидов после печени. Здесь происходят реакции липолиза и липогенеза.
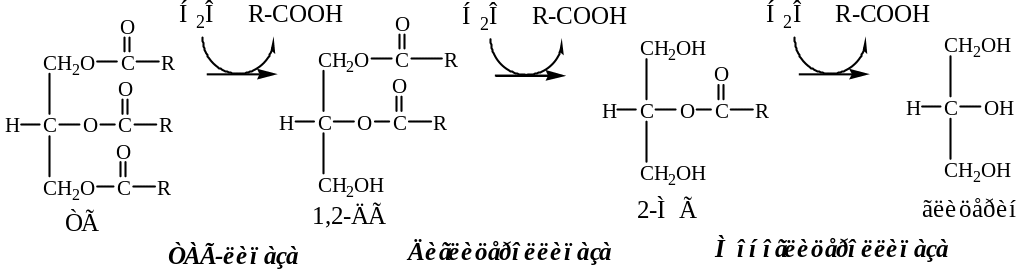
Липогенез. В жировой ткани синтез липидов идет в абсорбтивный период по глицерофосфатному пути. Процесс стимулируется инсулином.
Этапы липогенеза:
1. Под действием инсулина на рибосомах стимулируется синтез ЛПЛ.
2. ЛПЛ выходит из адипоцита и фиксируется на поверхности стенки капилляра с помощью гепарансульфата.
3. ЛПЛ гидролизует ТГ в составе липопротеинов
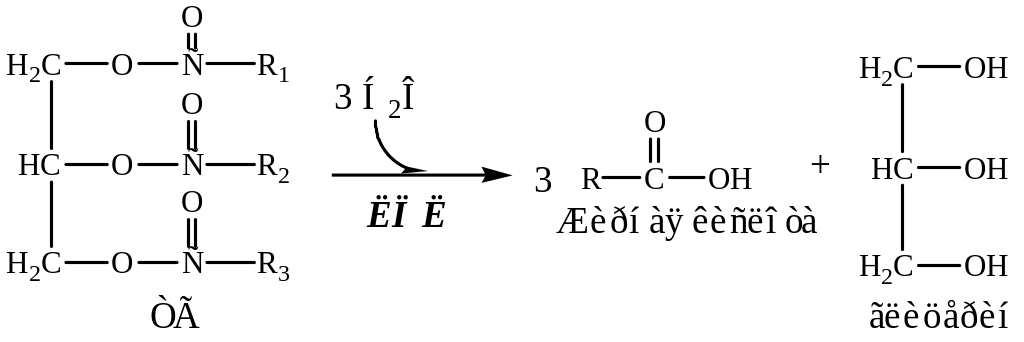
4. Образовавшийся глицерин уноситься кровью в печень.
5. Жирные кислоты из крови транспортируются в адипоцит.
6. Кроме поступающих из вне экзогенных жирных кислот, в адипоците синтезируются жирные кислоты из глюкозы. Процесс стимулируется инсулином.
7. Жирные кислоты в адипоците под действием Ацил-КоА синтетазы превращаются в Ацил-КоА.
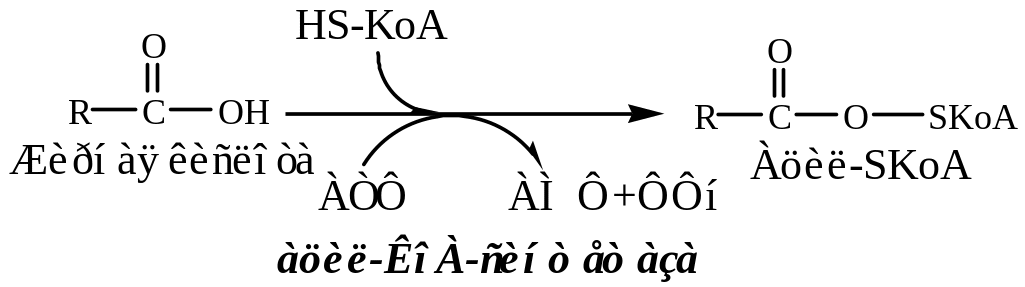
7. Глюкоза поступает в адипоцит с участием ГЛЮТ-4 (активатор инсулин).
8. В адипоците глюкоза вступает в гликолиз с образованием ФДА (активатор инсулин).
9. В цитоплазме ФДА восстанавливается глицерол-ф ДГ до глицерофосфата:
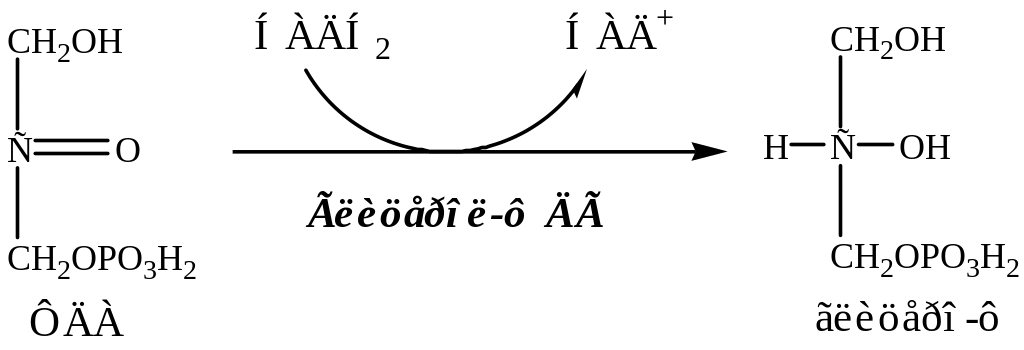
Так как в жировой ткани нет глицерокиназы, глицерофосфат образуется только из глюкозы (не может из глицерина).
10. В митохондриях глицерофосфат под действием глицеролфосфат ацилтрансферазы превращается в лизофосфатид:

11. В митохондриях лизофосфатид под действием лизофосфатид ацилтрансферазы превращается в фосфатид:

11. Фосфатид под действием фосфотидат фосфогидролазы превращается в 1,2-ДГ:

12. 1,2-ДГ под действием ацилтрансферазы превращается в ТГ:

13. Молекулы ТГ объединяются в крупные жировые капли.
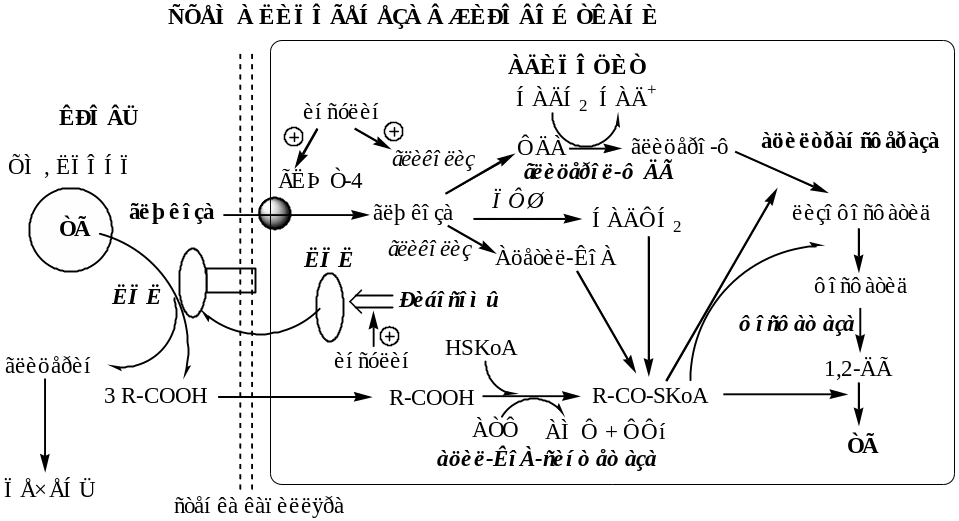
20.Механизм липолиза в белой жировой ткани: реакции, регуляция, значение
2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.
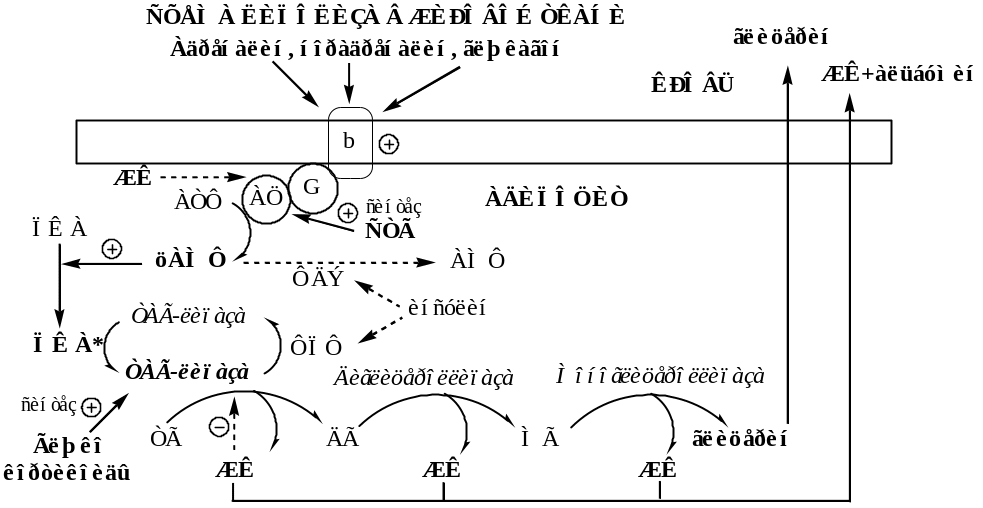
21.Механизмы регуляции липолиза – схема: роль СНС и ПСНС, их b- и a- адренорецепторов, гормонов адреналина, норадреналина, глюкокортикоидов, СТГ, Т3,Т4, инсулина и их внутриклеточных посредников, значение.
См схему Липолиза в жировой ткани!!!(выше)
Система, осуществляющая регуляцию липолиза в жировой клеточке, состоит из рецепторов на плазматической мембране клеток (альфа2-,бета2-и бетаЗ-адренорецепторы), G-белков, и гормон-чувствительной липазы. Альфа2-адренорецепторы ингибируют липолиз, а бета-рацепторы в комплексе с G-белками активируют липолиз. Регуляция липолиза осуществляется через аденилатциклазный механизм - при активации рецепторов происходит перевоплощение аденозинмонофосфата (АМФ) в повторяющийся аденозинмонофосфат (цАМФО, который, в свою очередь, активирует гормон-чувствительную липазу. Позже фермент фосфодиэстераза снова переводит цАМФ в АМФ, заблокируя таким образом липолиз
Мембрана жировой клетки обладает двумя типами рецепторов гормонов. С первыми взаимодействуют катехоламины (адреналин или норадреналин), усиливающие образование цАМФ, а значит, и липолиз, со вторыми взаимодействует инсулин, который снижает концентрацию цАМФ, противодействуя активации аденилатциклазы стимулирующими гормонами, и угнетает таким образом, липолиз. При сахарном диабете инсулиновая недостаточность приводит к бесконтрольной стимуляции внутриклеточной липазы и высвобождению большого количества ЖК из депо. Снижение концентрации глюкозы в крови при голодании подавляет выработку инсулина поджелудочной железой, что вносит свой вклад в усиление липолиза. Мозг постоянно нуждается в глюкозе, которая должна поступать с кровью. При снижении концентрации глюкозы в крови ниже критического уровня могут возникнуть тяжелые необратимые нарушения мозговых функций. При продолжительном голодании (запаса гликогена хватает ненадолго) мозг вынужден переключаться на использование бета-гидроксибутирата, образующегося в печени из жирных кислот. Тем самым запасы жира в организме позволяют сохранить в организме белки мышц, которые используются как последний источник глюкозы при голодании.)
Воздействие стресса (охлаждение, эмоции, нагрузки) мобилизует СЖК из депо либо за счет усиления секреции адреналина надпочечниками, либо способствуя высвобождению норадпреналина при стимуляции симпатических нервов, иннервирующих жировую ткань. Аденилатциклазную активность также повышают: глюкагон, адренокортикотропный гормон (АКТГ), гормон роста, гормоны щитовидной железы, вазопрессин. Тормозят липолиз: салицилаты, никотиновая кислота, РНК.
Такова биохимия углеводно-жирового обмена, эволюционно приспособленная для оптимального использования любых пищевых ресурсов. Преобдодание тех или иных процессов зависит от состава пищи, состояния и потребностей индивидуума и качественного и количественного характера запасов.
22.b-Окисление жирных кислот: кратко - история вопроса, суть процесса, современные представления, значение, тканевые и возрастные особенности.
Механизм окисления печенью жирных кислот был выяснен в начале нашего века. Он сводится к так называемому b-окислению. Окисление жирных кислот происходит до 2-го углеродного атома (b-атома). Получается более короткая жирная кислота и уксусная кислота, которая потом превращается в ацетоуксусную. Ацетоуксусная кислота превращается в ацетон, а новая b-окисленная кислота подвергается окислению с большим трудом. И ацетон и b-окисленная кислота объединяют под одним названием «кетоновые тела». Процесс окисления жирных кислот является важнейшим источником энергии в организме человека. Его значение для энергообеспечения различных органов и тканей (в первую очередь скелетных мышц и миокарда) выше, чем окислительного фосфорилирования глюкозы
Установлено, что окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза.
В 1904 г. Ф. Кнооп (F. Кnоор) выдвинул гипотезу бета-окисления жирных кислот на основании опытов по скармливанию собакам различных жирных кислот, в которых один атом водорода в концевой метильной группе (омега-углеродного атома) был замещен радикалом (С6Н5-).
Ф. Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в бета-положении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют чётное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов, проходит через стадию масляной кислоты.
После очередного бета-окисления масляная кислота становится ацетоуксусной. Последняя затем гидроли-зуется до двух молекул уксусной кислоты. Теория бета-окисления жирных кислот, предложенная Ф. Кноопом, в значительной мере послужила основой современных представлений о механизме окисления жирных кислот.