

3. Анатомічна будова кореня
Анатомічна будова кореня (мал. 2). Корінь, як і інші органи рослини, має клітинну будову. Різні його ділянки складаються з неоднакових клітин, що утворюють зони кореня. Це добре видно на молодих коренях цибулі, квасолі, соняшнику, пшениці та інших рослин.
На поздовжньому розрізі головні, додаткові й бічні корені мають подібну будову і в них можна виділити такі зони: зона росту з кореневим чохликом, зона розтягування (власне росту) і початок диференціації клітин, всисна і провідна зони.
Зона росту займає верхівку кореня завдовжки 2—3 мм. Це зона клітин, які активно діляться, меристема кореня.
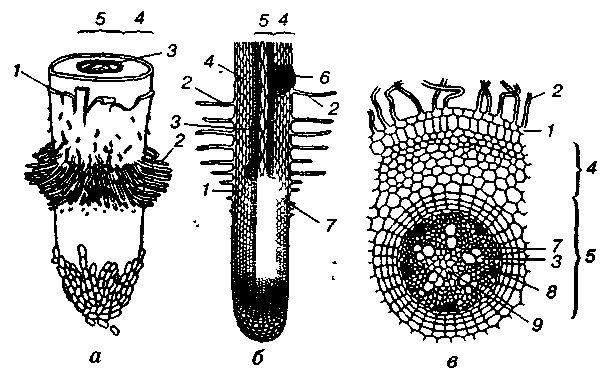
Мал. 2. Зони кореня і його первинна будова:
а — загальний вигляд; б — поздовжній розріз; в — поперечний розріз у зоні кореневих волосків: / — зона росту з кореневим чохликом; // — зона розтягування і початку диференціації клітин; /// — всисна зона; IV — провідна зона; 1 — волосконосний шар (епіблема, ризодерма); 2 — кореневі волоски; 3 — ксилема; 4 — кора кореня; 5 — центральний циліндр; 6 — початок росту бічного кореня; 7 — перицикл; 8 — флоема; 9 — серцевина
Усі тканини кореня виникають з цієї твірної тканини. Зона росту вкрита кореневим чохликом, який захищає верхівку кореня від пошкоджень під час просування кореня в ґрунті. Клітини кореневого чохлика мають підвищений тургор. Вони живуть недовго, поступово відмирають і злущуються. Замість відмерлих клітин постійно утворюються нові за рахунок зони поділу, яку прикриває кореневий чохлик (його немає у водяних рослин).
У зоні розтягування клітини ростуть, видовжуються і стають циліндричними. У них з'являються великі вакуолі. Сукупний ріст клітин цієї зони створює силу, завдяки якій корінь заглиблюється в грунт. Цьому сприяють "якірні" властивості кореневих волосків наступної зони кореня. Ця зона також невелика, всього кілька міліметрів. У верхній її частині клітини починають спеціалізуватися, і в зоні всисання повністю перетворюються на судини, трахеїди та інші види клітин кореня.
Всисна зона (завдовжки від кількох міліметрів до 1— 6 см) характеризується наявністю кореневих волосків — видовжених на 0,2—1,0 см виростів зовнішніх клітин кореня. Ядро клітини переходить у кореневий волосок і зазвичай розміщується в його верхівці. Завдяки великій кількості волосків (кілька сотень на 1 мм2) у рослин всисна поверхня збільшується в десятки разів. Наприклад, загальна площа всисної зони кореневої системи озимої пшениці може перевершувати площу надземної частини в 130 разів і становити 100 000 м2 на 1 га.
Кореневі волоски недовговічні, вони живуть 10—20 діб, а потім відмирають і злущуються. Нові волоски утворюються в процесі росту верхівки кореня в довжину. З ростом кореня в глибину переміщується і зона кореневих волосків.
За різних умов на коренях навіть однієї й тієї самої рослини кореневі волоски розвиваються неоднаково. Так, у сухому грунті вони розвиваються інтенсивніше, ніж у вологому. Коли вологи багато, вони не розвиваються зовсім. У болотяних або водяних рослин кореневих волосків зазвичай не буває, проте вони утворюються, коли ці рослини переходять на грунтові умови зростання. У епіфітних рослин, які мають повітряні корені, кореневі волоски розвиваються тільки тоді, коли корінь стикається з твердим субстратом. У деяких рослин роль кореневих волосків виконують гіфи гриба, з яким вони вступають у симбіоз.
Провідна зона, або зона бічних коренів (зона галуження), становить більшу частину кореня, вона розміщена над кореневими волосками і досягає кореневої шийки (місця переходу кореня в стебло). У цій зоні утворюються провідні судини і бічні корені. Провідна зона — це посередник між всисною зоною кореня і надземною частиною рослин.
У кореня розрізняють первинну і вторинну будову. Первинну будову мають молоді корені. В одних рослин така будова зберігається упродовж усього життя (більшість однодольних і незначна частина дводольних), а в більшості рослин первинна будова кореня змінюється на вторинну. Первинну будову мають корені всіх рослин у зоні кореневих волосків. На поперечному розрізі (див. мал. 2, в) добре помітні дві відокремлені частини: центральний циліндр, в якому є радіальний пучок, і периферична частина, що утворює кору кореня з кореневими волосками.
Кора кореня складається з ризодерми і первинної кори. Ризодерма (епіблема) — це первинна покривна тканина, клітини якої утворюють кореневі волоски. З ростом кореня клітини ризодерми відмирають, і покривною тканиною кореня стає екзодерма (за збереження первинної будови) або перидерма (за вторинної будови). Під ризодермою розміщена первинна кора кореня. Вона складається з паренхімних клітин, між якими е міжклітинники. Зовнішній шар клітин (екзодерма), що розміщений під ризодермою, складається з великих живих клітин. У них відкладаються крохмаль та інші поживні речовини. Ці клітини виконують захисну функцію і здатні пропускати воду та мінеральні солі від кореневих волосків до центрального циліндра. Після відмирання клітин ризодерми екзодерма перетворюється на покривну тканину.
Від центрального циліндра кора кореня відокремлена одним шаром мертвих клітин ендодерми. Внутрішні стінки цих клітин потовщені, скорковілі, не пропускають води і газів. Між мертвими клітинами ендодерми розміщені живі пропускні клітини, вони тонкостінні, розташовані навпроти судин центрального циліндра і легко пропускають розчини речовин до центрального циліндра.
Центральний циліндр займає середню частину кореня і складається з різних тканин. У периферичній частині його є перицикл, що складається з одного ряду тонкостінних клітин. Клітини перициклу (вторинна твірна тканина) періодично діляться і дають початок бічним кореням (звідси — коренетвірний шар), камбію, паренхімі кореня, додатковим брунькам коренепаросткових рослин. Основу центрального циліндра (усередині перициклу) становить паренхімна тканина, в якій радіальне розміщений судинний пучок кореня, що складається з ксилеми і флоеми. Судини ксилеми утворюють промені, що йдуть від периферії до центра, їх зазвичай буває три — п'ять, зрідка — близько 20. Між променями ксилеми розміщені групи клітин флоеми.
У більшості рослин (дводольних і голонасінних) первинна будова кореня зберігається недовго і переходить у вторинну будову. Така перебудова пов'язана з утворенням на певному етапі їхнього розвитку (після появи перших листків) у центральному циліндрі кореня вторинної меристеми — камбію. За рахунок клітин камбію утворюються вторинні елементи ксилеми і флоеми. У дерев і кущів вторинна ксилема і флоема наростають кільцями, тому будова кореня подібна до будови стебла. Первинна кора та ендодерма поступово відмирають і злущуються, а з перициклу утворюється перидерма. Нові шари перидерми закладаються у глибших шарах вторинної флоеми. Так поступово виникає вторинна будова кореня.
Внутрішня будова стебла деревної рослини. Особливістю будови стебла дводольної деревної рослини є його щорічне потовщення завдяки життєдіяльності камбію — вторинної твірної тканини. Внутрішню будову стебла деревної рослини добре видно на поперечному розрізі трирічної гілки липи.
У центрі розрізу розміщені великі клітини серцевини з тонкими оболонками, в них можуть відкладатися поживні речовини. У багатьох дерев серцевина пухка, може відмирати й утворювати у стовбурі порожнину — дупло. Серцевину оточують три (за числом років зрізаної гілки) концентричних шари деревини приблизно однакової товщини.
Це свідчить про подібні умови росту гілки упродовж трьох років. Деревина складається з судин, клітин паренхіми і механічної тканини. Численні клітини механічної тканини надають стеблу міцності і захищають великі судини від стискання сусідніми клітинами. До периферії зовнішнього кільця прилягає тонкий шар клітин вторинної твірної тканини — камбію. Зовні від камбію розміщується шар кори, вкритий корковою тканиною і відмерлою шкіркою.
У корі є різні тканини: провідна, механічна, основна. Провідна тканина представлена ситоподібними трубками, механічна — луб'яними волокнами, які надають стеблу міцності й гнучкості. Основна тканина виповнює проміжки між цими тканинами і може нагромаджувати поживні речовини.
Зовні кора вкрита корковою тканиною, яка складається з мертвих клітин, на її поверхні знаходяться клітини відмираючої шкірки. Шкірка і корок захищають стебло від впливу несприятливих умов, не пропускають вони і повітря. У корковому шарі більшості дерев і кущів є спеціальні отвори, крізь які надходить повітря, необхідне для дихання рослини, — сочевички, які мають вигляд маленьких горбиків на поверхні стебла. Сочевички утворені великими клітинами основної тканини з великими міжклітинниками.
Потовщення стебла відбувається за рахунок періодичної діяльності камбію. Клітини камбію постійно діють у період вегетації рослини і можуть функціонувати упродовж кількох років. Особливо інтенсивно розмножуються клітини камбію навесні, тому кора з пагонів легко знімається. Схематично процес росту клітин камбію можна уявити так: клітина камбію росте і ділиться. З двох нових клітин внутрішня спеціалізується і перетворюється на клітину деревини, що прилягає до старих її елементів, а клітина камбію, відсунута назовні, зберігає здатність до подальшого поділу (тобто залишається камбіальною). Після поділу цієї клітини зовнішня дочірня клітина починає перетворюватися на постійну клітину кори, а внутрішня дочірня залишається камбіальною. Ця камбіальна клітина знову утворює клітину деревини, а далі — кори. Отже, камбій відкладає клітини безперервно: всередину — клітини, з яких будується деревина, назовні — клітини, з яких будується кора. Нові клітини деревини наростають назовні, а клітини кори — із середини. При цьому камбій весь час кільцеподібне обхоплює деревину.
Як правило, клітин деревини утворюється значно більше, ніж клітин кори, тому деревина займає значнішу частину на поперечному розрізі стебла. Клітини деревини і кори, які утворилися з камбію, можуть розростатися як у поздовжньому, так і в поперечному напрямку.
Ріст клітини камбію припиняється з настанням зимового періоду, а навесні поновлюється. Лише у посушливих районах (саванах) рослини влітку втрачають листя і вступають у літній період спокою. За умов вологих тропіків дія камбію не припиняється, тому рослини там ростуть постійно.
На поперечному розрізі трирічного пагона липи видно, що деревина має чітко виражені шари. Це річні кільця, які чітко виділяються внаслідок того, що деревина, яка утворилась у різні пори року, має різні колір, блиск і щільність. Весняна деревина, яка утворилася після відновлення діяльності камбію, складається з великих клітин з тонкими оболонками. Вона зазвичай багатша на судини та трахеїди, тому здається пухкішою і забарвлена в світліший колір. Осіння деревина складається з більш товстостінних, але вужчих волокон і замість елементів, які проводять воду, містить більше механічної тканини, тому здається темнішою. Внаслідок відмінності у структурі пізня деревина функціонує переважно як механічна тканина, а весняна проводить воду з розчиненими мінеральними солями. Перехід від весняної деревини до осінньої відбувається поступово, а перехід від осінньої до весняної завжди раптовий.
Ширина річних кілець залежить від виду дерева, його віку (на початку життя рослини утворюються ширші кільця, з віком вони поступово вужчають), від умов середовища. За шириною кілець можна прочитати історію умов життя дерева.
Видозміни пагонів. Крім основної функції, про яку вже йшлося, пагони виконують низку додаткових функцій, пов'язаних з їх видозмінами (метаморфозами), які виникли в процесі пристосувальної еволюції. Це вусики — довгі тонкі пагони з редукованими листками. Вусиками (виноград, огірок та інші рослини з лазячими або повзучими стеблами) рослини прикріплюються до різних предметів. Колючки — вкорочені пагони без листків, що захищають рослину від поїдання тваринами (терен, дика яблуня, груша). Вусики і колючки — це видозміни надземного пагона. Вони розміщуються в пазухах листків або на листковому вузлі навпроти листка, що свідчить про стеблове походження їх.
Крім видозмін надземного пагона у рослин є видозміни і підземного: кореневище, бульба, цибулина, які виконують функції нагромадження поживних речовин і вегетативного розмноження.
Кореневище — це видозмінений пагін з невеликими лускатими листками буруватого кольору і бруньками.
Кореневище зовні схоже на корінь, але відрізняється наявністю рудиментарних листків, бічних і верхівкових бруньок і відсутністю кореневого чохлика. За формою воно може бути довгим і тонким (пирій, осока) або коротким і товстим (щавель, іриси). Щороку з верхівкової бруньки виростає надземний пагін. Під час обробітку грунту кореневище може бути подрібнене, тоді кожна його частина з брунькою дасть нову рослину. Особливо швидко відбувається вегетативне розмноження рослин з розгалуженими кореневищами (пирій).
Бульба — це потовщена верхівка підземного стебла — столона. Найбільш відомими є бульби картоплі. Про стеблове їх походження свідчить наявність і спіральне розміщення бруньок-вічок. Хлорофілу бульби не мають, але на сонці зеленіють. Бульби можуть бути і надземними, наприклад потовщене стебло капусти кольрабі. Такі бульби мають зелений колір.
Цибулина — дуже вкорочений підземний пагін з видозміненими листками. Цибулини бувають кулясті, яйцеподібні, видовжені тощо. Стеблова частина цибулини займає незначну зону, її називають денцем. На нижній поверхні денця розміщені численні додаткові корені, на верхній — видозмінені м'ясисті листки (луски), що щільно прилягають один до одного і мають запас поживних речовин. Зовнішні листки (луски) виконують захисну функцію, тому стають шкірястими. У пазухах деяких соковитих лусок є бруньки, з яких розвиваються або дочірні цибулини (дітки), або надземні зелені листки і квітконосні "стрілки".
Цибулини формуються у багатьох лілійних рослин: цибулі, лілії, тюльпана тощо. У деяких лілійних можуть бути надземні цибулини. Зазвичай вони утворюються у суцвіттях (у дикої цибулі, часнику), але можуть міститися і в пазухах листків.