

Движение глаз и активация корковых зрительных нейронов
Как уже упоминалось в начале этой главы, зрительное восприятие тесно связано с движениями глаз и головы, изменяющими направление взгляда. Поэтому неудивительно наличие множества различных соединений между корковыми зрительными областями и глазодвигательными центрами в стволе мозга. Так, сигналы из области V2 идут в верхние холмики четверохолмия, которые управляют контролирующими направление взгляда нейронами в ПМРФ и РФСМ (с. 238). Экстраст-
1 )
Речь идет об узнавании знакомых лиц
(близких, родственников
и т.д.), а не лиц «вообще». Последняя
способность
при прогнозировании сохраняется.- Прим.
пер.
)
Речь идет об узнавании знакомых лиц
(близких, родственников
и т.д.), а не лиц «вообще». Последняя
способность
при прогнозировании сохраняется.- Прим.
пер.
262 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
|
Рис. 11.31. Средняя частота импульсации (ось ординат) двух нейронов «лицеспецифичной» корковой области в глубине верхней височной борозды (см. рис. 11.30). Бодрствующим животным предъявлялись стимулы, показанные в нижнем ряду (2,5 с каждый красные горизонтальные полоски). Нейрон 1 дает максимальный ответ на стимул угловым размером 5°, соответствующий профилю обезьяны, а нейрон 2-на лицо «анфас». Если закрыть горизонтальную часть лица на уровне глаз, реакция нейрона 2 меняется незначительно. С другой стороны, предъявление щетки вызывает слабый его ответ (по [40] с изменениями) |

|
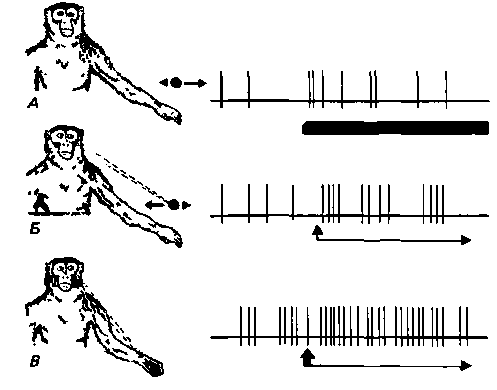
Рис. 11.32. Схема ответа нейрона поля 7 бодрствующего макака-резуса. А. Объект (например, орех) находися в пределах досягаемости и движется; движение возбуждает нейрон (частота его импульсации увеличивается). Б. Животное смотрит на объект; частота импульсации возрастает еще больше. В. Когда животное хватает объект, возбуждение нейрона достигает максимума [47] |
риарные зрительные области связаны с префронтальными корковыми зонами, которые отвечают за произвольные движения глаз (поле 8) и за координацию движений кисти и рта. В свою очередь, эти области, подобно полю 7 теменной коры (см. ниже), соединены с глазодвигательными центрами ствола мозга и - обратными связями -с затылочной зрительной корой. Путь из сетчатки через нейроны верхних холмиков (с. 257) важен для афферентной передачи зрительной информации о движении в поле 7. Нейроны верхних холмиков посылают сигналы в подушку таламуса, откуда они идут к экстрастриарным корковым зрительным областям и полю 7 теменной коры. Эти связи также способствуют интеграции движений взгляда и зрительного восприятия.
Нейронное отображение окружающего пространства
Координаты и пространственная организация объектов в окружающем мире кажутся нам неизменными, даже когда их изображения на сетчатке смещаются при активных движениях глаз, головы и тела. Более того, по крайней мере в той части внешнего пространства, которая находится в пределах непосредственной досягаемости, предметы имеют мультимодальные свойства: чашка воспринимается как таковая, когда мы смотрим на нее, когда прикасаемся к ней и когда из нее пьем. Обширные поражения теменной коры головного
мозга человека нарушают восприятие предметов в пространстве или полностью препятствуют ему. Как правило, этот дефицит ограничен контралатеральной месту поражения половиной пространства (одностороннее зрительное игнорирование)1). Обычно он сопровождается нарушением произвольных движений глаз в контралатеральную сторону. Функцию теменных полей коры изучали с помощью микроэлектродов, вживленных в мозг обезьяны. В поле 7 здесь находятся нейроны, возбуждаемые как зрительными, так и тактильными стимулами. Их активность частично зависит от движений глаз, хотя на нее влияют также интерес и та часть окружающего пространства, на которое животное обращает внимание (рис. 11.32). Очевидно, что в поле 7 отображение окружающего не зависит от ретинотопической проекции. Зрительные области подушки, посылающие сигналы к полю 7, также участвуют в обеспечении этих сложных функций [5, 39, 48, 58, 59].
1 ) У правшей встречается преимущественно синдром «игнорирования левой половины пространства» при поражениях теменной доли правого полушария. Многие авторы специально выделяют его как показатель поражения именно правого полушария.- Прим. пер.