

11.4. Нейрофизиология и психофизика восприятия света и темноты
Когда в ясный день на солнце набегает облако, мы замечаем уменьшение освещенности, к которому вскоре адаптируемся. Даже при стократном изменении силы света воспринимаемые относительные светлота и цвета окружающих объектов изменяются весьма незначительно. В условиях фотопического зрения восприятие контраста и цвета сравнительно независимо от средней освещенности. Другими словами, вид предмета определяется отражательной способностью его поверхностей, а не абсолютным количеством отраженного от них света. Данный механизм важен для понимания удивительной неизменности видимого нами мира.
Если при постоянном уровне адаптации освещается определенная область сетчатки, наблюдается приблизительно логарифмическая зависимость между воспринимаемой субъективной светлотой светового пятна и его измеренной яркостью (уравнения 9, 10). С помощью микроэлектродных записей показано, что такой же зависимостью описывается частота импульсации нейронов с оп-центрами (рис. 11.17, В). У нейронов с off-центрами она также приблизительно соответствует логарифмической функции предшествующего снижения стимуляции при «выключении света». В целом отмечается линейная зависимость между субъективной
Глава 11. Зрение 253
|
Рис. 11.20. Функциональная организация рецептивных полей ганглиозных клеток сетчатки млекопитающих. При анализе рецептивных полей небольшие пятна света (показаны белым) проецировались либо на центр, либо на периферию РП. Световые стимулы вызывают разный ответ у нейронов с on- и off-центром. Когда обе части РП освещены одновременно, возбуждающий и тормозный процессы, связанные с освещением центра и периферии, суммируются. Однако преобладает ответ, вызываемый стимуляцией центра РП |
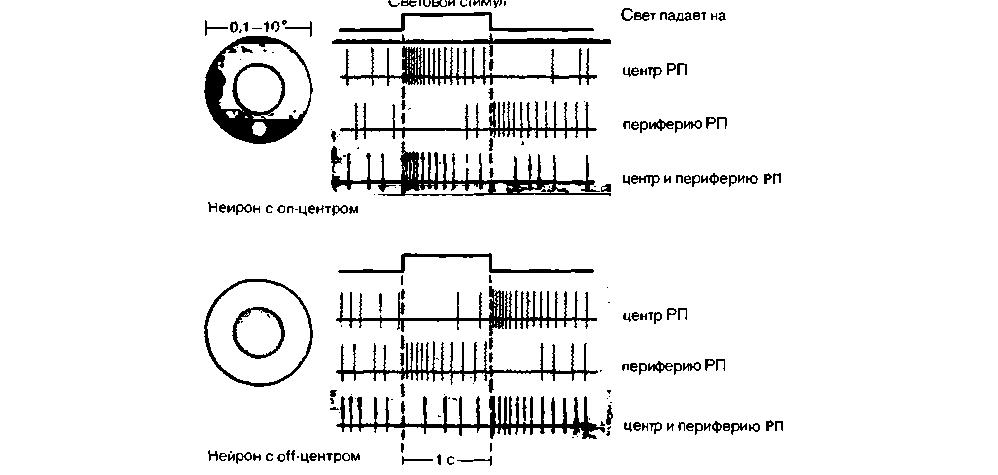
светлотой и средней частотой импульсации нейронов оп-системы и между субъективной темнотой и средней частотой импульсации нейронов offсистемы. Зная это простое правило корреляции нейронной активности и восприятия, можно объяснить целый ряд элементарных зрительных феноменов.
Нейрофизиологическая основа одновременного контраста
Рассмотрим первый пример корреляции между зрительным восприятием и возбуждением on- и off-нейронов. Серое пятно на белом фоне кажется темнее, чем на черном (рис. 11.21). Вдоль границы между темным и светлым наблюдаются полосы Маха (пограничный контраст): ближайшая к ней светлая часть кажется светлее, а ближайшая темная-темнее, чем более удаленные от границы. Исходя из функциональной организации РП ганглиозных клеток сетчатки, можно предложить следующее объяснение этого простого примера одновременного контраста (рис. 11.22). Одновременный контраст - важный механизм, обеспечивающий частичную функциональную компенсацию физиологических недостатков диоптрического аппарата, описанных на с. 244-245, и в результате повышающий остроту зрения [35].
Изменение остроты зрения и организации рецептивных полей при изменениях окружающей освещенности
Каждый знает по опыту, что острота зрения снижается от центра поля зрения к его периферии и что она гораздо хуже при скотопическом зрении, чем при фотопическом. В фотопических условиях острота зрения снижается при уменьшении средней освещенности.
Вы сможете прочесть такой мелкий шрифт, только если будете смотреть прямо на него, а страница книги будет достаточно хорошо освещена.
Зависимость контрастности восприятия и остроты зрения от средней освещенности можно объяснить реорганизацией РП ганглиозных клеток сетчатки (рис. 11.23). Очевидно, что порог прямого возбуждения (за счет центра РП) соответствует меньшей силе света, чем для латерального торможения, однако эффективность последнего при росте освещенности увеличивается быстрее, чем у возбуждения. Если пространственная организация РП соответствует суммации возбуждения и торможения (рис. 11.23), диаметр функционального центра РП уменьшается при возрастании средней освещенности. Далее, исходя из соотношения числа рецепторов и ганглиозных клеток (в среднем примерно 125:1), очевидна весьма выраженная конвергенция сигналов из слоя рецепторов в слое ганглиозных клеток. Однако в зоне центральной ямки она значительно меньше. Соотношение конвергенции и латерального торможения в этой области сетчатки при оптимальных условиях освещения позволяет достичь остроты зрения, близкой к теоретическому пределу, определяю-
254 ЧАСТЬ ΠΙ. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
|
Рис. 11.21. Рисунки, иллюстрирующие явление одновременного яркостного контраста |

|
Рис. 11.22. Схема механизма одновременного контраста. А В. Возбуждение нейрона с оп-центром в зависимости от положения в РП границы темного и светлого. Максимальная активация соответствует случаю Б, когда граница между темным и светлым совпадает с границей между центром и периферией РП. Числа соответствуют относительной амплитуде возбуждения (без учета спонтанной активности нейрона). Г. Зависимость возбуждения нейронов зрительной системы от положения в РП границы темного и светлого (ось абсцисс). Эти упрощенные кривые отражают ответы нейронов сетчатки с on- и off-центрами, а также «контрастовых» нейронов ЛКТ с оп-центрами |
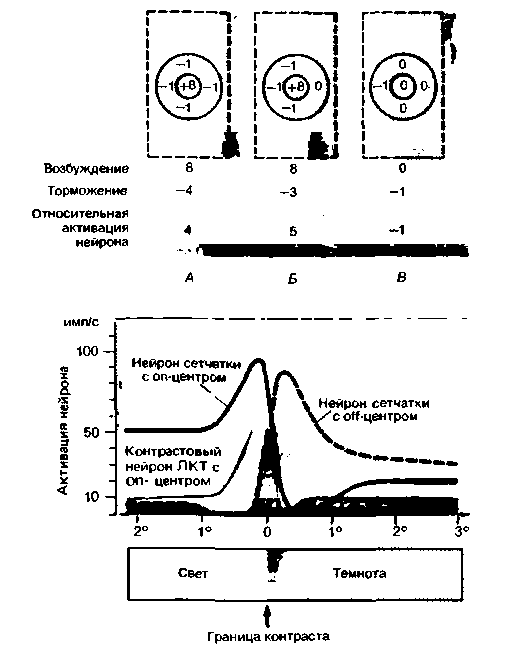
|
Рис. 11.23. Пространственная организация рецептивного поля ганглиозной клетки on-типа, объясняемая суперпозицией пространственных распределений возбуждающего и тормозного процессов. У возбуждения порог ниже, чем у торможения. По мере роста освещенности в надпороговой области, если торможение увеличивается быстрее возбуждения, центр РП будет функционально уменьшаться. При очень ярком (слепящем) свете преобладает торможение: световая реакция ганглиозной клетки оп-типа тормозится и зрительное восприятие формы нарушается. При оптимальном уровне световой адаптации размеры центра РП минимальны, следовательно, острота зрения максимальна (Grüsser, Fortschr. Ophthalmol., 1983, 80, 502). Ц - центр РП; Π периферия РП |
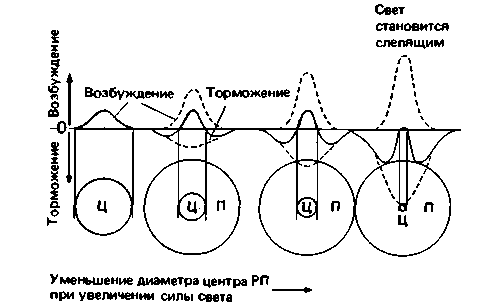
щемуся пространственной мозаикой популяции колбочек 0,5', что примерно соответствует диаметру наружных сегментов находящихся здесь фоторецепторов.
Снижение остроты зрения - не единственное отрицательное последствие неадекватного освещения читаемой книги или рабочего места. Из-за кажущейся нерезкости изображения рефлекторно включаются механизмы аккомодации (с. 244), которые, разумеется, оказываются в данном случае неэффективными. Когда система аккомодации непрерывно напряжена, легко может возникнуть головная боль. Поэтому важно обеспечить надлежащее освещение рабочего места и избегать при этом источников света с неустойчивой яркостью.