

Роговично-сетчаточный потенциал и электроретинограмма (эрг)
С помощью макроэлектродов от всего глаза можно отводить два функционально различных типа электрических потенциалов: постоянный между роговицей и более отрицательной сетчаткой - и электроретинограмму (ЭРГ). Первый обусловлен главным образом разностью потенциалов между склеральной стороной клеток пигментного эпителия и внутренними сегментами фоторецепторов. Другими словами, он отражает суммарный электрический ток, проходящий через плазматические мембраны пигментных клеток и фоторецепторов, поэтому меняется в зависимости от уровня адаптации сетчатки.
ЭРГ-это колебания отводимого от всего глаза напряжения, вызываемые увеличением или уменьшением количества падающего на сетчатку света. ЭРГ включает несколько «волн»: а, b, с и d (рис. 11.19). Волна а возникает, по-видимому, за счет суммации рецепторных потенциалов; более медленная b-волна отражает изменения, главным
Глава 11. Зрение 251
|
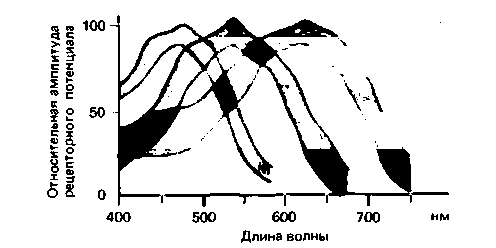
Рис. 11.18. Спектральная чувствительность рецепторных потенциалов сетчатки рыбы. Показаны средние значения (со стандартным отклонением) результатов для трех разных типов колбочек (по [61] с изменениями) |
|
Рис. 11.19. Электроретинограммы сетчатки человека (по Hanitzsch et al.. Vision Res. 1966, 6, 245, схематизировано) |
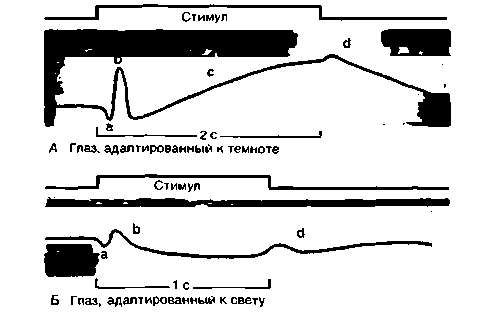
образом мембранных потенциалов биполярных и глиальных (мюллеровых) клеток, с-волна - мембранных потенциалов клеток пигментного эпителия при «включении света» (on-эффект), а d-волна— мембранных потенциалов фоторецепторов и биполярных клеток при «выключении света» (off-эффект). Если длительность светового стимула превышает 0,3 с, с-волна появляется еще в ходе его действия и off-эффект накладывается на нее. При коротких световых вспышках b- и d-волны совпадают [8, 10].
ЭРГ обращаемого узора регистрируется при рассматривании узора, светлые и темные участки которого периодически меняют яркость на противоположную (например, при обращении цвета светлых и темных шахматных полей на телеэкране). Средняя яркость такого стимула неизменна, поэтому ЭРГ в данном случае генерируется в основном ганглиозными клетками сетчатки, т.е. нейронными элементами, особенно сильно реагирующими на изменения
контраста и контрастные границы. Если ганглиозные клетки дегенерируют, ЭРГ обращаемого узора исчезает. Такой симптом в медицине обычно связан со слепотой или с резким ухудшением остроты зрения (см. с. 264).
Рецептивные поля нейронов сетчатки
Нейронная сеть сетчатки. Вторичный рецепторный потенциал передается через синаптические контакты от рецепторов биполярным и горизонтальным клеткам с помощью химического медиатора (см. с. 51). Рецептивные поля (РП) последних (см. с. 184) распространяются на несколько фоторецепторов. Если осветить фоторецепторы центра РП биполярных клеток оп-типа, мембрана этих биполяров деполяризуется (рис. 11.13), если периферии РП гиперполяризуется. Гиперполяризация осуществляется косвенно - за счет контактов между горизонтальными и биполярными клетками или же через петлю обратной связи между горизонтальными клетками и синаптическими пузырьками колбочек.
РП горизонтальных клеток, как правило, сравнительно велики. Функционально они связаны друг с другом. Некоторые горизонтальные клетки (L-muпа) гиперполяризуются при попадании света на их РП независимо от его спектрального состава; другие при освещении либо деполяризуются, либо гиперполяризуются в зависимости от того, какой части спектра соответствует стимул (цветоспецифичные красно-зеленые или сине-желтые горизонтальные клетки, с. 273).
Биполярные клетки off-типа гиперполяризуются при попадании светового стимула в центр их РП и деполяризуются, когда он падает на периферию РП (рис. 11.13). Сетчатка содержит примерно равные количества on- и off-биполяров. Функциональная организация их слоя определяется двумя важнейшими принципами обработки зрительной информации нейронами.
Возбуждение фоторецепторов фотонами передается в ЦНС по двум независимым нейронным «каналам»: через on-систему, активируемую, когда световая стимуляция в центре РП сильнее, чем на периферии или чем предыдущая стимуляция центра, и через off-систему, активируемую при уменьшении освещения.
Рецептивные поля многих биполяров организованы антагонистически, т.е. световой стимул вызывает их диаметрально противоположную реакцию в зависимости от того, попадает он на периферию или в центр РП. Такая пространственная организация РП обеспечивается двумя способами: конвергенцией сигналов от нескольких рецепторов на одной биполярной клетке и опосредованными тормозными сигналами с периферии РП, доходящими
252 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
до биполяров через горизонтальные клетки (латеральное торможение, с. 186).
Этот принцип конвергенции сигналов и латерального торможения действует и на следующем уровне обработки информации в сетчатке (ганглиозные и амакриновые клетки, рис. 11.13). Как правило, несколько on- («прямое» возбуждение) и off- («прямое» торможение) биполяров конвергируют на одну ганглиозную клетку с on-центром, однако значительно больше биполярных клеток косвенно связаны с последней посредством контакта с амакриновыми клетками формируя периферию ее с РП (латеральные on-торможение и off-возбуждение). Ганглиозные клетки с off-центром возбуждаются off-биполярами, прямо тормозятся оп-биполярами (центр РП), «латерально» тормозятся off-амакриновыми клетками и, наконец, латерально возбуждаются амакриновыми клетками, возбуждаемыми оп-биполярами [11, 17, 18, 33, 49].