

Глава 11. Зрение 249
|
Рис. 11.15. Результаты микроспектрофотометрических измерений спектров поглощения одиночных рецепторов сетчатки человека (образцы взяты во время операций). Кривые дифференциальные спектры (разница спектров поглощения до и после выцветания). Можно различить три типа колбочек (по Brown, Wald, Science, 1964, 144, 45, схематизировано) |

вых каналов мембраны рецептора; цГМФ заставляет их оставаться «открытыми». Тогда первичный фотохимический процесс состоит в восстановлении за несколько миллисекунд цГМФ за счет цепочки ферментативных реакций: натриевая проводимость уменьшается и возникает гиперполяризационный вторичный рецепторный потенциал. Предполагается, что при этом молекула родопсина, превратившись в метародопсин II за счет поглощения фотона, в крайне быстрой последовательности активирует множество молекул фермента («трансдуцина»), что приводит к начальному «усилению» первичного фотохимического процесса, поскольку «трансдуцин» управляет цГМФ [4, 5, 8, 34, 46, 53, 56а].
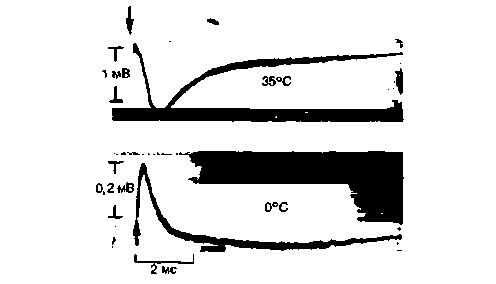
Первичный рецепторный потенциал фоторецепторов и фотохимические компоненты световой и темновой адаптации. Конформационное изменение молекулы родопсина генерирует с очень коротким латентным периодом (меньше 1 мс) электрический потенциал, называемый первичным (ранним) рецепторным потенциалом (РРП) (рис. 11.16). В нем несколько компонентов, которые можно выявить, постепенно снижая температуру сетчатки. Когда зрительный пигмент распадается на ретинол и опсин, происходит его ресинтез за счет цепи поглощающих энергию ферментативных реакций. Если на сетчатку падает постоянный поток фотонов, устанавливается динамическое равновесие между процессами выцветания зрительного пигмента под действием света и его регенерации с участием ферментов. Если сетчатки достигает меньше света, оно сдвигается в сторону более высокой концентрации родопсина. Это фотохимическая основа хорошо всем известной темновой адаптации глаза (см. с. 254, 255). В ходе нее амплитуда РРП, вызываемого световыми стимулами постоянной интенсивности, повышается, поскольку возрастает вероятность соударения падающих фотонов с молекулами невыцветшего родопсина [8].
Вторичный рецепторный потенциал. Если РРП генерируется за счет синхронных конформационных изменений молекул зрительного пигмента, то вторичный (поздний) рецепторный потенциал ПРП, не связанный с РРП непосредственно, - это изменение мембранного потенциала фоторецептора. В темноте он составляет от —25 до —40 мВ; под действием света происходит гиперполяризация. Амплитуда
|
Рис. 11.16. Ранний рецепторный потенциал фоторецепторов бурундука, записанный при двух различных температурах (по Pak, Ebrey: J. gen. Physiol., 1966, 49, с изменениями). Амплитуда различных его компонентов возрастает примерно пропорционально логарифму силы световой вспышки. Стрелкой показан момент вспышки |
этого рецепторного потенциала растет с увеличением интенсивности светового стимула (рис. 11.17), причем у палочек медленнее, чем у колбочек. Другими словами, временная инерция палочковой системы больше, чем у колбочковой (с. 256). Неодинаковая спектральная чувствительность вторичных рецепторных потенциалов различных типов колбочек (рис. 11.18) подтверждает результаты микроспектрофотометрии и говорит в пользу трихроматнческой (трехкомпонентной) теории цветового зрения (с. 271). т.е. присутствия трех типов колбочек с разными спектральными чувствительностями. Спектральная чувствительность рецепторных потенциалов палочек примерно соответствует спектру поглощения родопсина с максимумом около 500 нм.
У позвоночных рецепторный потенциал фоторецепторов в отличие от других рецепторов связан
250 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
|
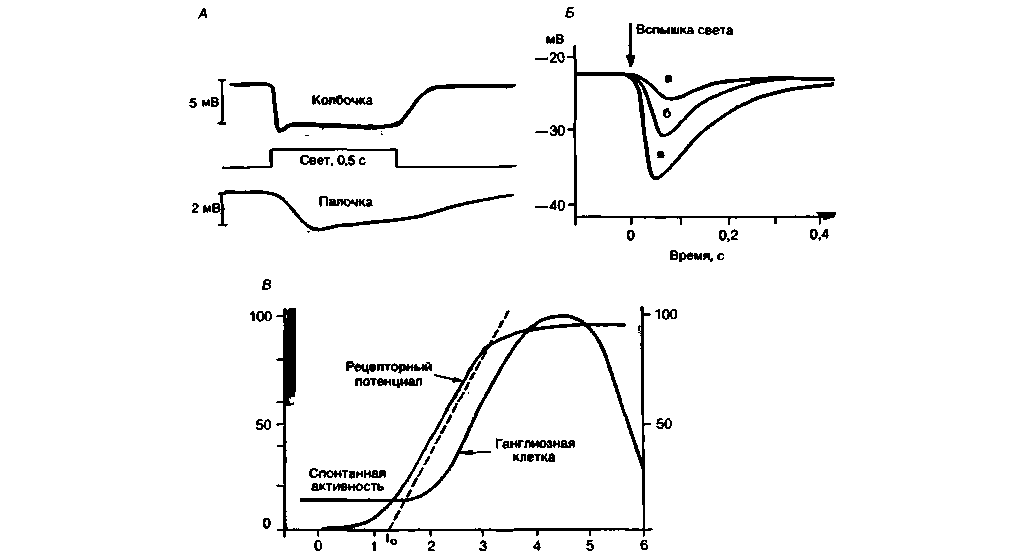
Рис. 11.17. Поздний рецепторный потенциал фоторецепторов сетчатки позвоночных. А. Внутриклеточная регистрация рецепторных потенциалов колбочки и палочки (схематично). Б. Рецепторный потенциал колбочки сетчатки черепахи в ответ на световые вспышки длительностью Юме с возрастающей интенсивностью (относительные интенсивности: а = 1. б = 4. в = 16). В. Кривая амплитуды рецепторного потенциала одиночной колбочки сетчатки черепахи. Относительная амплитуда (ось ординат) увеличивается пропорционально десятичному логарифму относительной интенсивности стимула. ls (ось абсцисс) в сравнительно узком диапазоне. Показана также схематическая кривая зависимости величины возбуждения ганглиозной клетки сетчатки с on-центром от силы света (Б, В: по Baylor. Fuortes J. Physiol. (Lond.) 1970. 207) |
не с деполяризацией, а с гиперполяризацией мембраны в ответ на адекватную стимуляцию (свет).
Амплитуда (А) вторичного рецепторного потенциала коррелирует с интенсивностью светового стимула (Is) (т.е. с числом фотонов, падающих в единицу времени на единицу площади) следующим образом (рис. 11 .17, Б, В):
![]() (9)
(9)
Эта гиперболическая функция («закон Геринга») в диапазоне промежуточных интенсивностей приближается к логарифмической, которая в общей сенсорной физиологии известна как закон ВебераФехнера (см. с. 188, 189):
![]() (10)
(10)
где I0- пороговая интенсивность стимула, зависящая от уровня адаптации. Константы а, к и к* в уравнениях (9) и (10) меняются вместе с длиной волны монохроматического света. Эта зависимость примерно соответствует кривым спектральной чувст-
вительности рецепторных потенциалов (рис. 11.18) [8, 36].