

Відділ Зелені водорості - Chlorophyta
Відділ є найчисленнішим серед водоростей. За даними різних авторів, він нараховує 20-25 тис. видів. Найдавніші рештки відомі з верхнього докембрію — протерозою (біля 1400 млн. років). Зелені водорості поширені в прісних водоймах, в морях та океанах, в наземних біотопах, на снігу та льоду.
Від зелених водоростей походять вищі рослини.
До складу фотосинтетичних пігментів зелених водоростей входять майже всі пігменти, властиві вищим рослинам: хлорофіли "а" та "Ь", всі відомі каротини (а, Р, у та є-каротин), ксантофіли лютеїно- вого ряду (лютеїн, зеаксантин, неоксантин, віолаксангин, антераксан- тин). Завдяки переважанню хлорофілів, більшість видів Chlorophyta забарвлені у зелений колір. В екстремальних умовах у деяких зелених водоростей "вмикається" механізм гіперсинтезу каротину, і тоді клітини набувають червоного забарвлення.
Основний продукт асиміляції - крохмаль, який завжди відкладається в хлоропласті, часто - навколо піреноїду. Додаткові асиміляти - олія та лейкозин. У сифонофіцієвих водоростей основним асимілятом може бути полісахарид інулін.
Клітини переважної більшості видів вкриті целюлозно-пектиновою оболонкою. Зрідка зустрічаються представники з голими клітинами.
амінопластів немає. Характерна особливість бриопсидальних - нияиність у життєвому циклі стефаноконтних зооспор. Розмножуються іірионсидальні фрагментацією талому, апланоспорами, стефано- міііі'ними зооспорами та статевим шляхом. Статевий процес - і е гсрогамія. Типовий представник порядку' - Вгуорлі.ч.
Рід бриопсис - Вгуоряіз поширений у морях тропічного,
убтропічного та помірного поясів.
Рис. 65. Вгуорхії. Загальний вигляд талома
Гаметофіти цієї водорості диференційовані на сланкі ризоїдальні та висхідні асиміляторні сифони. Висхідні сифони зазвичай пірчасто розгалужені і нагадують пташине пір’я. При утворенні гамет деякі бічні сифони відокремлюються перегородкою. З сифонів, забарвлених в інтенсивно зелений колір, утворюються жіночі гаметангії, з світліших, жовтуватих сифонів - чоловічі. Далі вміст гаметангіїв розпадається на численні дводжгутикові жіночі та чоловічі гамети, що різняться за розмірами. Через отвір в оболонці гамети виходять у зовнішнє середовище, де попарно копулюють, утворюючи планозиготу.
 Планозигота
осідає на субстрат, втрачає джгутики і
без періоду спокою починає проростати
у диплоїдний розгалужений ниткоподібний
спорофіт, що отримав назву стадії
ОегЬе<;іа. Розвиток стадії ОегЬеБІа
відбувається 2-4 місяці. Ядро у 20-40
разів збільшується у діаметрі, переходячи
зі стадії диплоїдного зиготичного ядра
у стадію первинного ядра. Далі у первинному
ядрі иідбувається серія мітотичних
поділів, дочірні ядра зменшуються у
діаметрі до 5-10 мкм. Останній поділ ядра
є редукційним. Після мейозу завершується
формування вторинних ядер, і спорофіт
холо- або еукарпічним шляхом дає початок
одноядерним стефаноконтним гаплоїдним
зооспорам. Ці зооспори проростають або
у нові і аметофіти, які складаються з
сифонів пірчастої будови, або у нові і
порофіти стадії ОегЬеяіа. Цікаво, що за
певних умов планозигота може безпосередньо
проростати у гаметофіт, не проходячи
стадію І ктЬсзіа. Проте, процеси, що
відбуваються з ядром при цьому шляху
розвитку, залишаються недослідженими.
Планозигота
осідає на субстрат, втрачає джгутики і
без періоду спокою починає проростати
у диплоїдний розгалужений ниткоподібний
спорофіт, що отримав назву стадії
ОегЬе<;іа. Розвиток стадії ОегЬеБІа
відбувається 2-4 місяці. Ядро у 20-40
разів збільшується у діаметрі, переходячи
зі стадії диплоїдного зиготичного ядра
у стадію первинного ядра. Далі у первинному
ядрі иідбувається серія мітотичних
поділів, дочірні ядра зменшуються у
діаметрі до 5-10 мкм. Останній поділ ядра
є редукційним. Після мейозу завершується
формування вторинних ядер, і спорофіт
холо- або еукарпічним шляхом дає початок
одноядерним стефаноконтним гаплоїдним
зооспорам. Ці зооспори проростають або
у нові і аметофіти, які складаються з
сифонів пірчастої будови, або у нові і
порофіти стадії ОегЬеяіа. Цікаво, що за
певних умов планозигота може безпосередньо
проростати у гаметофіт, не проходячи
стадію І ктЬсзіа. Проте, процеси, що
відбуваються з ядром при цьому шляху
розвитку, залишаються недослідженими.
представників мають вигляд нерозгалужених ниток або розгалужених кущиків. У деяких морських родів таломи представлені системою розгалужених міхурів, або складаються з центральної вісі та з'єднаних
з нею бічних сегментів, або набувають псевдопаренхіматозного вигляду.
Життєвий цикл гаплодиплофазний, з ізоморфною зміною поколінь. Клітинні оболонки целюлозні, позбавлені пектинових речовин. Вегетативні клітини мають багато ядер, пристінний сітчастий хлоропласт або велику кількість дрібних дископодібних хлоропластів, з'єднаних між собою дуже тонкими тяжами; у хлоропластах розташовуються дрібні піреноїди. Кожний піреноїд поділенй на дві напівсфери, навколо яких відкладається суцільна напівсферична платівка крохмалю. Мітози у різних ядрах однієї клітини відбуваються синхронно, проте цитокінез та каріокінез між собою не зкоординовані. Монадні клітини позбавлені субмікроскопічних лусочок; зооспори переважно чотириджгутикові, гамети — дводжгутикові. У кладофоральних виявлено специфічний ксантофіл - сифоноксантин. Характерні представники - СІасІорИога та Югігосіопіит.
Рід кладофора - СІскІорИога є провідним родом порядку. Таломи кладофори кущикоподібні, складаються з багатоядерних клітин, що утворюють системи розгалужених ризоїдальних та висхідних ниток. Клітини звичайно вкригі товстими шаруватими оболонками, до яких прикріплюються численні епіфітні організми. В цитоплазмі окремої клітини міститься пристінний сітчастий хлоропласт з багатьма пірено'іцами та численні ядра. В живому стані ядра не помітні в світловий мікроскоп, тому щоб їх роздивитися рекомендується забарвити препарати ацетокарміном. Водорість розмножується фрагментацією таломів, зооспорами, акінетами та статевим шляхом. Статевий процес - ізогамія.
При нестатевому розмноженні у верхівкових клітинах спорофіту відбувається мейоз, після якого утворюється велика кількість чотири- джгутикових зооспор. Через отвір в оболонці спорангію зооспори виходять у зовнішнє середовище і проростають у гаметофіт. Гаметофіти кладофори є роздільностатевими. Чоловічі та жіночі гамети морфологічно не відрізняються між собою, проте різняться фізіологічно та за ультратонкою будовою апікальної частини клітини. Так, жіночі гамети на верхівці під плазмалемою мають спеціальний ковпачок, який при копуляції приймає участь в утворенні кон'югаційного каналу. Хоча
(іудь-яка вегетативна клітина висхідної частини талому здатна перетворитись у гаметангій, проте процес утворення гамет, як правило, починається у верхівкових клітинах талому. Через отвір в ні »олонці гамети звільняються і копулюють з гаметами особин іншого і і/ігевого знаку. Після копуляції зигота осідає на субстрат і без періоду спокою починає проростати у новий спорофіт.
Гамети кладофори (а також і деяких інших кладофоральних) ідатні проростати партеногенетично, даючи початок як спорофітам, гак і гаметофітам. Ця особливість зумовлює існування в межах роду досить різноманітних варіантів конкретних життєвих циклів, які, проте, є похідними від основного, гаплодиплофазного циклу зі гноричною редукцією.
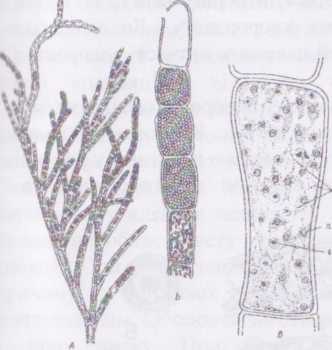 Рис.
68.
СІаііорЬога.
Рис.
68.
СІаііорЬога.
А - частина нитки з зооспорангіями (темні клітини); Б - зооспорангії;
В - багатоядерна клітина: п - піре- ноїди; я - ядра; х — хроматофор.
Найпоширенішим видом морів помірних та теплих широт є Сіасіоріюга \agabunda. У прісних водоймах найчастіше трапляється С. glomerata. Морфологічно ці два види не відрізняються між собою, але перший має гашюдиплофазний життєвий цикл зі зміною поколінь, тоді як у другого статевий процес відсутній, а нестатеве розмноження нідбувається дводжгутиковими зооспорами. Спеціальні дослідження показали, що зооспори С. glomerata за походженням є гаметами, що и тратили здатність до копуляції, проте зберігли здатність до партено- гонетичного розвитку. Таким чином, вегетативні таломи найпоширенішого прісноводного виду роду - С. glomerata - це гаплоїдні гаметофіти.
Клас Кон’югати - Соп|і^аіорЬусеае
Об'єднує одноклітинні та нитчасті зелені водорості у яких відсутні у життєвому циклі джгутикові стадії. Нестатевого розмноження іа участю зооспор в них немає, а статевий процесе - кон'югація -
полягає у злитті амебоїдних протопластів звичайних вегетативних клітин. Досить поширене вегетативне розмноження. Для більшості представників класу притаманний центральний осьовий.хроматофор з піреноїдами.
Кон’югати - прісноводні водорості,але зустрічаються серед них грунтові форми та та такі,що мешкають у солонуватих водах.
Клас включає чотири порядки, найважливіші серед них - Zygnematales та Безтісііаіез.
Порядок Зигнематальні - 7.уепста(а!еу
Об'єднує водорості з нитчастим типом морфологічної структури тіла, у яких джгутикові стадії та центріолі відсутні. Життєвий цикл гаплофазний, без чергування поколінь, з зиготичною редукцією.
На роди зигнематальні поділяються, в першу чергу, за типом морфологічної структури та формою хлоропласту. До найбільш поширених зигнемагальних водоростей належать нитчасті водорості з родів Бріго&га, Іу&іета та Mougeotia.
Рід спірогіра - Зріго^у га. Нитки цієї водорості складаються з витягнугих або коротких циліндричних клітин. Клітинна оболонка целюлозна, зовні вкрита слизовим шаром, виразно помітним у розчині туші, а також на дотик. Поперечні клітинні перегородки часто утворюють різноманітні зморшки.
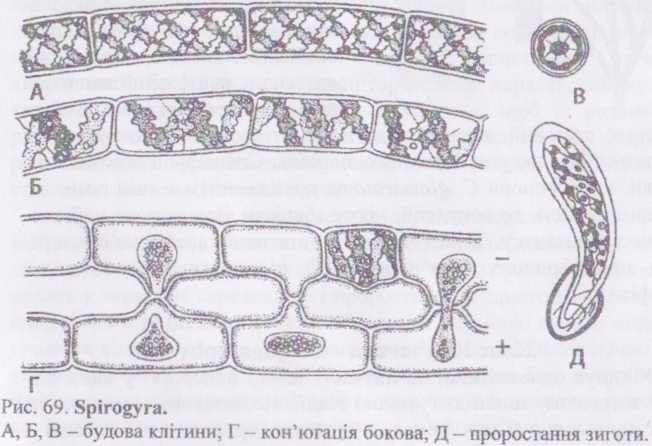
У кожній клітині міститься від одного до кількох (здебільшого І І) стрічковидних спіральних хлоропластів з піреноїдами. Краї
ч поропластів нерівні, інколи розсічені. Центр клітини займає велика ішкуоля з клітинним соком. Ядро велике, з добре помітним ядерцем. У чистини видів воно кулясте, в інших має лінзовидну форму. Ядро "підвішене" в центрі вакуолі в цитоплазматичному мішечку, від якого на різні боки до периферії
Вегетативне розмноження у спірогіри здійснюється фрагментацією ниток внаслідок відмирання окремих інтеркаляриих клітин. Ці фрагменти, і навіть окремі клітини, можуть проростати у нові нитки.
Статевий процес - коньюгація. Розрізняють драбинчасту та бокову коньюгацію. Більш поширена драбинчаста коньюгація, яка відбувається між кпітинами двох ниток. В цьому випадку нитки розміщуються паралельно одна одній, і об'єднуються спільним слизом. Далі від клітин протилежних ниток назустріч одна одній починають рости копуляційні відростки, які стикаються своїми кінцями. Оболонки в зоні контакту розчиняються і утворюється кон'югаційний канал. По ньому протопласт однієї клітин поступово проштовхується в іншу. Там протопласти та ядра зливаються і утворюється зигота. Оскільки поведінка клітин при коньюгації відрізняється, то сприймаючу клітину називають жіночою, а віддаючу - чоловічою, іигота виробляє товсту тришарову оболонку, і переходить у стан спокою. При проростанні диплоїдне ядро зиготи редукційно ділиться, причому з чотирьох гаплоїдних ядер лише одне залишається життєздатним. Оболонка зиготи розривається, і протопласт проростає у нову нитку. При бічній коньюгації копуляційні відростки утворюються між сусідніми клітинами однієї нитки.
Види роду 5ріго&га поширені у стоячих або повільно текучих кодах: заводях, старицях річок, ставках, болотах, канавах, калюжах. Часто спірогіра утворює великі маси яскраво-зеленого жабуриння, яке слизьке на дотик.
Рід зигнема - Zygnema Види цього роду за зовнішнім виглядом жабуриння нагадують спірогіру, але при мікроскопії відрізняються (’»удовою хлоропластів: в кожній клітині їх два, вони розміщені в центрі, мають зірчасту форму. В кожному хлоропласті добре помітний один великий піреноїд.
Рис.
70. Ху%пепіа.
Будова
клітини.
Рід мужоція - Мои^еоНа. Види цього роду також утворюють темно-зелене слизьке жабуриння, і часто суцільно вкривають поверхню водойм. Від зігнеми та спірогіри мужоція відрізняється, в першу чергу, наявністю в кожній клітині лише одного хлоропласта, який має вигляд платівки з двома або кількома піренощами і розміщується по вісі клітини.
Рис.
71. Мої^еоПа. Будова клітини.
Порядок Десмідіальні - Рсатііііаісц
Включає кокоідні водорості, у яких джгутикові стадії та центріолі відсутні, статевий процес - кон'югація, мітоз напіввідкритий. Клітинні оболонки переважно скульптуровані, обов'язково мають простий або складний поровий апарат. У примітивніших десмідіальних клітини не диференційовані на напівклігини, у досконаліших клітина поділена на дві напівклітини, що з'єднуються між собою перешийком, в якому розташовується ядро. Подібно до зигнематальних, життєвий цикл гаплофазний, без чергування поколінь, з зиготичною редукцією.
Найбільш поширеними та багатими у видовому відношенні представниками порядку є Сіозіегіит та Сойтагіит.
Рід клостеріум - СІомегіит. Водорість має поодинокі, веретеноподібно зігнуті клітини. Клітинна оболонка гладенька або злегка штрихувата і містить багато дрібних та дві великі нори. Останні знаходяться на полюсах клітини і виконують функції локомоторних органів - через них клітина виділяє слиз. Виділення слизу або спричинює повільний реактивний рух, або слиз утворює швидкоростучі слизові ніжки, за допомогою яких клітина змінює своє положення у просторі. При слабкому освітленні у клітин спостерігається позитивним фототаксис, при інтенсивному - негативний. Хлоропласти нагадують осьові конуси з радіальними поздовжніми ребрами; по осі \ поропласту розташовується кілька піреноїдів. Па полюсах клітин утворюються вакуолі з кристалами гіпсу.
Водорість розмножується поділом клітин надвоє та внаслідок і глтевого процесу - кон'югації. Зигота, що утворюється внаслідок кон'югації, переходить у стан спокою, по завершенні якого відбувається мейоз. Далі два з чотирьох гаплоїдних ядер дегенерують, і зигота проростає двома проростками.
І’ис.
72. Склепиш. А - вигляд клітини збоку; Б
- поперечний розріз клітини; В - виділення
мас слизу з кінців клітини; Г - кон’югація;
Д,Е - кон’югація з утворенням подвійних
зигот.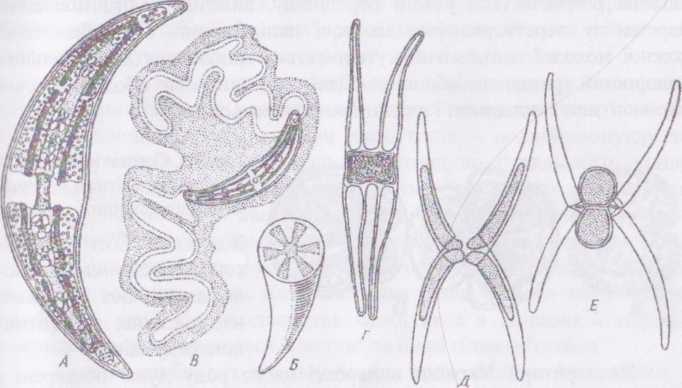
Рід налічує більше двохсот видів, серед яких біля 70 знайдені на території України. Види роду широко розповсюджені в різноманітних прісних водоймах, рідше зустрічаються на вологих скелях та мохах.
Рід космаріум - Сохтагіит. До цього роду належать кокоїдні мікроскопічні прісноводні водорості, що мають клітини з двох і и метричних напівклітин. Останні з'єднуються між собою широким ибо вузьким перешийком. Напівклітини не розсічені на лопаті, хоча їх форма може бути дуже різноманітною (наприклад, округлою, сіііпсоїдною, майже квадратною, кутастою тощо).
Клітинні оболонки зазвичай скультуровані бородавками, шипами, шипиками, нерідко просякнуті солями заліза. Клітини мають складний поровий апарат, що складається з багатьох дрібних "та двох- чотирьох великих слизових пор, розташованих на полюсах клітин. Слизові пори забезпечують рух клітини, механізм якого такий самий, як і у Сіоьіепит. Кожна напівклітина містить один або два хлоропласти з піреноїдами, а в зоні перешийку - одне ядро.
Розмножується космаріум вегетативним поділом або внаслідок кон’югації. При вегетативному поділі у перешийку за способом фрагмопласту утворюється клітинна перегородка. Кожна молода клітина розростається у зоні перешийку, внаслідок чого половини перешийку перетворюються на нові напівклітини. Під оболонкою кожної молодої напівклітини утворюється типова для десмідіальних водоростей тришарова оболонка. Далі зовнішній шар оболонки нової напівклітини скидається, і клітини роз'єднуються.
 Рис.
73. Созтагіиія: а
- вигляд клітини
спереді, б — ПОДІЛ КЛІТИНИ.
Рис.
73. Созтагіиія: а
- вигляд клітини
спереді, б — ПОДІЛ КЛІТИНИ.
Життєвий цикл водоростей роду Созтагіит гап-
лофазний, без чергування поколінь, з зиготич- ною редукцією.
На території України водорості цього роду дуже поширені і різноманітні за видовим складом. Більшість видів належить до (3- мезосапробів і мешкає у прісних водоймах різного типу (річках, озерах, ставках, калюжах). Проте, найчастіше види космаріуму зустрічаються у сфагнових болотах. Частіше за інші трапляються С. гепі/огте - космаріум ниркоподібний, С. ругат 'иіаіит - космаріум пірамідальний, С. та^агИі/егит - космаріум перламутровий та С. еаеІаіит - космаріум вирізаний.
Клас Харофіцієві - СЬагорЬусеае
Об'єднує багатоклітинні прісноводні та морські водорості з особливою, так званою харофітною морфологічною структурою тіла та багатоклітинними статевими органами.
Клас представлений одним порядком та трьома родинами.
Порядок Харальні - СЬагаІез
Об'єднує водорості з гетеротрихальним типом морфологічної
іруктури тіла, які мають макроскопічні членисто-мутовчасті таломи, /їжіутикові стадії, що представлені сперматозоїдами, оогамний статевий процес, багатоклітинні жіночі статеві органи. Харальні подорості мешкають у бентосі озер та приморських солонуватоводних пиманів. Характерними представниками порядку є СИага та Шеііа.
Рід хара - СИага. Спорофіти харальних виглядають як розгалужені кущики, що складаються з прямостоячих опірних ниток необмеженого росту - т.зв. "стебел", бічних мутовчасто розміщених асиміляторних ниток обмеженого росту - "листків", та ниток-ризоїдів. Кожна нитка ("стебло", "листок", ризоїд) диференційована на клітини вузлів та міжвузлів, що правильно чергуються одна з одною.
Вузли та міжвузля утворюються внаслідок поділу апікальної клітини. Вона ділиться у поперечній до вісі нитки площині, по черзі відокремлюючи від себе клітини двох типів - подвійно-опуклу та нодвійно-увігнуту. Подвійно-опукла клітина до подальшого поділу нездатна - вона лише росте та розтягується, поступово диференціюючись у велетенську багатоядерну клітину міжвузля. Подвійно- увігнута клітина здатна до подальших поділів. Ці поділи відбуваються у радіальних напрямках у поздовжній до осі клітини площині. Дочірні клітини утворюють вузел. Кожна клітина вузла, в свою чергу, здатна до поділу, який відбувається так само, як і в апікальній клітині, внаслідок чого утворюються "листки" та бічні гілки - "стебла".
З клітин стеблових вузлів у деяких представників (наприклад, у СИага) розвиваються стеблова кора та прилистники, з листових вузлів спорангії, що дають початок чоловічим або жіночим гаметофітам. На ризоїдальних вузлах утворюються вивідкові бруньки (бульбочки), що
органами вегетативного розмноження.
Клітини вкриті оболонками, зазвичай просякуються вапном. Центр клітини зайнятий великою вакуолею з клітинним соком. І (.итоплазма з органелами утворює тонкий периферичний шар.
Клітини міжвузлів багатоядерні. їх ядра мають характерну лопатеву форму, крім звичайних ядерних структур (хроматину, ядерець, прорибосом) вони містять пучки мікротрубочок і діляться амітотично. Клітини вузлів, навпаки, одноядерні, сферичні, їх поділ відбувається шляхом мітозу. Всі клітини спорофіту позбавлені центріолей. В клітинах чоловічих гаметофітів, навпаки, центріолі є і
