Клас Улотриксові - іЛоігісЬорИусеае
Об'єднує переважно морські бентосні та перифітонні водорості, хоча в межах класу представлені також і прісноводні та аерофітні форми. Майже всі ульвофіцієві водорості мають багатоклітинні таломи. Переважаючі типи морфологічної структури тіла - нитчастий, гетеротрихальний та сифонокладальний.
Клітини улотриксових завжди вкриті целюлозно-пектиновою або целюлозною оболонкою. Монадні клітини завжди голі, чотири- джгутикові або дводжгутикові, представлені зооспорами та гаметами.
До класу входить біля 80 родів, що об'єднують більш ніж 1000
видів.
Система класу. Ознаки, за якими клас поділяють на порядки, пов'язані з типами морфологічної структури тіла, життєвими циклами, ультратонкою будовою зооспор та гамет, а також з особливостями поширення. До класу входить шість порядків, з яких нижче розглядаються чотири - ІЛоІхісЬаІез, ІЛуаІея, СЬаеІіорЬогаІез, Оес^опіаІеБ.
Порядок Улотрихальні - ЦіоІгісЬаІса
Об'єднує морські та прісноводні водорості з нитчастим та гетеротрихальним типом структури. Життєвий цикл гаплодиплофазний, з гетероморфною зміною поколінь. Спорофіт представлений диплоїдною одноклітинною рослиною, відомою під назвою кодіолюм (Сосііоіит). Клітинні оболонки містять велику кількість пектинових речовин і здатні ослизнюватися. Монадні клітини голі, але на плазмалемі є шар субмікроскопічних органічних
м уі очок. Зооспори переважно чотириджгутикові, гамети (іподжгутикові. Типовий представник - ІЛоікгіх.
Рід улотрикс - иіоі'ггіх Більшу частину життя водорість іі|н»водить у стані гаметоспорофіту, який має вигляд довгих мсрозгалужених однорядних ниток. У кожної клітини нитки (за нпнятком базальної) є пристінний хлоропласт у вигляді незамкненого кільця, кілька пірено'ідів, одне ядро; центр клітини зайнятий вакуолею І клітинним соком. Всі клітини здатні до поділу, що обумовлює постійне наростання талому в довжину, а також утворення І ч* продуктивних клітин. Базальна клітина, за допомогою якої водорість прикріплюється до субстрату, відрізняється від звичайних негстативних клітин нитки: вона витягнута у короткий ризоїд, не щічна до поділу, має редукований хлоропласт.
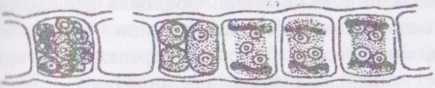 Рис.
57.
Рис.
57.
Улотрикс (иісгіїтх). Нитка з зооспорами, які розвиваються в деяких клітинах.
Вегетативне розмноження відбувається фрагментацією нитки. І Іри нестатевому розмноженні у будь-якій клітині утворюються 2-4 (8) чоіириджгугикових зооспор. Зооспороутворення розпочинається в .піікальній частині нитки і просувається до її основи. Зрілі зооспори ииходять назовні у слизовому міхурі через боковий отвір клітинної і іінки. Міхур швидко розчиняється. Після певного періоду руху зооспора зупиняється, відкидає один за одним усі чотири джгутики, прикріплюється боком до субстрату і проростає в нову нитку.
Статевий процес ізогамний. Будь-яка клітина нитки здатна утворити велику кількість (8-64) дводжгутикових ізогамет, які для популяції виходять у зовнішнє середовище. Нитки ІЛоікгіх є роздільностатевими, оскільки, незважаючи на морфологічну схожість гамет, одні нитки продукують лише фізіологічно жіночі гамети ("плюс"-гамети), інші нитки - фізіологічно чоловічі (”мінус"-гамети). І Іісля копуляції зигота деякий час зберігає джгутики і рухається, тобто перебуває у стані планозиготи. Далі вона зупиняється, втягує
І ж гутики, виробляє товсту оболонку, і переходить у стан спокою. ГІО його завершенні зигота проростає у булавоподібний одноклітинний > порофіт (стадія СосЗіоІит): вона формує трубковидний випин, куди переходить цитоплазма та ядро; даті випин здувається, набуваючи
59
грушоподібної форми; потім диплоїдне ядро спорофіту редукційно ділиться і спорофіт утворює 4-8 чотириджгутикових зооспор. Останні через отвір у клітинній оболонці виходять назовні у слизовому міхурі; слиз швидко розчиняється, і зооспори звільняються. Після деякого періду активного руху зооспори осідають на субстрат і проростають, даючи початок новим нитчастим гаметоспорофітам.
Види роду поширені переважно у морях, де зустрічаються у супраліторальних ваннах або в обростаннях літоральної зони. Кілька видів досить часто зустрічаються у прісних водоймах - переважно в холодних гірських річках та струмках, а також у весняний сезон у річках та озерах помірного поясу.
Порядок Ульвальні - Ціуаіев
Включає майже виключно морські водорості з гетеротрихальною або примітивною тканинною будовою. Житгєвий цикл гапло- дигшофазний, з ізоморфною зміною поколінь. Оболонки целюлозно- пектинові, здатні до ослизнення. Монадні клітини ульвальних позбавлені субмікроскопічних лусочок. Зооспори перевалено чотири- джгутикові, гамети дводжгутикові. Характерні представники - ІІН-а та ЕШеготогрка.
Рід ульва, або зелений морський салат - \Jlva. Спорофіти та гаметофіти водорості морфологічно однакові, і мають вигляд двошарової платівки з гофрованими краями, довжина якої складає кілька сантиметрів. Кожна клітина містить одне ядро, пристінний пластинчастий хлоропласт з одним-кількома піреноїдами.
А
- зовнішній вигляд талому; Б
- поперечний розріз таллома.
Цикл розвитку
ульви.
Таломи гаметофітів розвиваються з чотириджгутикових зооспор,
норофітів - з щойно утворених зигот, які зберігають джгугики гамет, іоЬто із планозигот. Розвиток пластин спорофітів та гаметофітів йідбувається із зооспор або планозигот однаковим способом: монадна і її піна прикріплюється до субстрату, втрачає джгутики і ділиться і початку у поперечній площині, утворюючи однорядну нерозгалужену норвинну нитку, яка нагадує талом улотриксу. Далі клітини нитки шляться повздовжньо-радіально, утворюючи одношарову трубку з іюрожниною в центрі. Стінки трубки змикаються, і крайові клітини продовжують ділитися, внаслідок чого утворюється двошарова
на іївка зі складчастими краями.
Хоча таломи спорофітів та гаметофітів морфологічно однакові, проте плоїдність їх різна, і тому зооспори і гамети ніколи не утворюються на одному й тому ж таломі. Зооспори утворюються шляхом послідовного поділу протопласту будь-якої клітини на чотири — вісім кнітин, причому перший поділ є редукційним, і. таким чином, спорофіти є диплоїдними, зооспори - гаплоїдними.
Статеве розмноження відбувається за допомогою дводжгутико- ііих гамет. Статевий процес, залежно від виду, може бути ізогамним ибо гетерогамним. Дня ульви характерна гетероталічність - гаметофіти иідрізняються в статевому відношенні: одні продукують гамети зі ніаком інші - зі знаком Копуляція відбувається лише між іамстами різних статевих знаків. Якщо статевий процес ізогамний, "+" і а таломи розрізниш не вдається. У випадку гетерогамії таломи різних статевих знаків розпізнаються навіть неозброєним оком, за моарвленням плодючих ділянок. Так, у чоловічих таломів ці ділянки -кош уваті, у жіночих - темно-зелені.
Зигота, що утворюється внаслідок копуляції гамет, містить дип- моїдне ядро і без періоду спокою одразу проростає в диплоїдний (іюрофіт. Таким чином, у ульви спостерігається правильне чергування гюморфних поколінь: диплоїдного спорофіту та гаплоїдного і амстофіту.
Види роду иі\-а мешкають в морях теплого та помірного поясів, юкрема, в Чорному морі. Водорість є їстівною, і в деяких країнах миніть введена в аквакультуру і вирощується на спеціалізованих морських фермах.
Рід ентероморфа, або кишечниця - Епіеготогрка. У цього (иіизького до ульви роду таломи видовжено-пластинчасті або трубчас
ті. За типом життєвого циклу, будовою клітин та розмноженням цей рід подібний до \Jlxa. Відміни полягають у морфогенез! талому, ню й відбивається на зовнішньому вигляді водорості. Рис. 59. ЕпІегошогрЬа.
 А
- зовнішній вигляд;
А
- зовнішній вигляд;
Б - поперечний розріз стінки талома.
Зокрема, у ентєроморфк талом розвивається з зооспори або планозиготи через стадію однорядної первинної нитки, далі - одношарової трубки з порожниною в центрі. Як тільки стінки трубки у верхівковій частині змикаються, морфологічні трансформації талому, за винятком його лінійного збільшення у довжину, припиняються. Як наслідок, молода водорість має вигляд кишки (звідси її народна назва - кишечниця), а доросла у нижній частині представлена одношаровою трубкою з порожниною, у верхній - двошаровою видовженою платівкою.
Кишечниця поширена переважно в морях, де часто зустрічається разом з ульвою. Проте, деякі види роду добре переносять значне опріснення води, завдяки чому піднімаються у гирла річок і навіть розвиваються у прісних континентальних водоймах.
Порядок Хетофоральні - СЬаеіорЬогаіея
Об'єднує багатоклітинні водорості з нитчастим та гетеро- трихальним типом морфологічної структури тіла. Каркас клітинної оболонки хетофоральних утворений фібрилами кристалічно впорядкованої целюлози. Таломи хетофоральних представляють фізіологічно цілісні структури, оскільки клітини у нитках сполучаються між собою за допомогою плазмадесм.
Монадні клітини - зооспори та гамети - голі. Зооспори мають чотири джгутики, гамети дводжгутикові. Всі хетофоральні водорості мешкають у перифітоні прісних континентальних водойм.
В межах порядку простежується поступове ускладнення плану будови тіла: від нитчастих нерозгалужених водоростей до гетеротри- хальних представників, що складно диференційовані у морфологічному та функціональному відношенні. Цей ряд добре простежується на прикладі родів ІІгопета, БИ^еосІопіит, ОгарагпаШа.
Рід уронема — игопета має талом у вигляді простої нерозгалуженої нитки, що складається з морфологічно та функціонально однакових клітин (за винятком базальної та апікальної). Кожна клітина одноядерна, має пристінний хлоропласт у вигляді незамкненого кільця, в якому розташовується один (рідше - два-чотири) ггіреноїди. Базальна клітина утворює диск, за допомогою якого водорість прикріплюється до підводних субстратів. Апікальна клітина має звужену та загострену верхівку. Розмноження відбувається за допомогою чотириджгу- тикових зооспор, або апланоспорами.
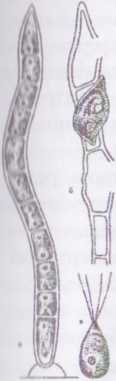 Рис.
60. игопета:
Рис.
60. игопета:
а) вегетативна нитка; б) утворення зооспор; в) зооспора.
Рід стигеоклоніум - $Ґі%еосІопіит має гетеротрихальні таломи, що складаються з розгалужених висхідних та сланких ниток. Перші ідійснюють функції фотосинтезу та розмноження, за допомогою других водорість прикріплюється до субстрату. Висхідні нитки на нсрхівці звужені, і досить часто закінчуються одно- або багатоклітинними тонкими безбарвними волосками - хетами. Рясність галуження
і її кількість хет залежить від концентрації азоту у середовищі: якщо у йоді міститься велика кількість азоту, галуження зустрічається зрідка, і
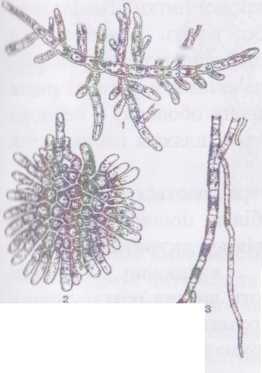 хети
майже відсутні (в цьому випадку ~ водорість
нагадує игопета),
і навпаки.
хети
майже відсутні (в цьому випадку ~ водорість
нагадує игопета),
і навпаки.
Сі /Г Нестатеве розмноження відбувається
чотириджгутиковими зооспорами та ап- ланоспорами, статевий процес - ізогамія, здійснюється дводжгутиковими гаметами. Цікаво, що гамети здатні проростати партеногенетично, і, таким чином, виконувати також функції зооспор. Життєвий цикл гаплофазний, із зиготичною редукцією і без зміни поколінь.
Рис. 61. Будова Б^еосіопіиш:
2 - будова сланкої частини слані, 3 - ризоїди
Рід драпарнальдія - ВгаратаМіа. Водорість макроскопічна, має вигляд тендітного кущика, вкритого м'яким прозорим слизом, може досягати 10-20 см у висоту. Таломи ИгаратаШіа диференційовані на три системи ниток — сланкі, висхідні опірні та асимілятори. Сланкі нитки виконують функції прикріплення до субстрату, асимілятори - фотосинтезу та розмноження. Опірні нитки підносять над субстратом всю масу асиміляторів.
Клітини опірних ниток мають товсті клітинні оболонки, редукований хлоропласт у вигляді поясковидної, по краю зазубреної стрічки, іноді продірявленої або майже сітчастої, з численними піре- ноїдами. Нитки-асимілятори короткі, рясно розгалужені, на верхівках часто закінчуються волоском (хетою). Клітини ниток-асиміляторів містять великий хлоропласт з кількома піреноїдами і здатні утворювати зооспори та гамети. Клітини сланких ниток безбарвні.
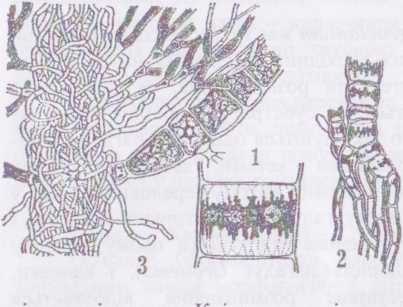 Рис.
62 ОгарагпаИіа:
Рис.
62 ОгарагпаИіа:
- клітина головної осі;
- ризоїди;
- частина слані Вгарагааісііорзіз.
Нестатеве розмноження здійснюється мікро- і макрозооспорами, розвиток яких відбувається одночасно в усіх клітинах окремої нитки-асимілятора або навіть одночасно в усіх асиміляторах, крім того, клітини асиміляторів можуть утворювати апланоспори, які іноді залишаються з'єднаними у вигляді рядів або купок. Апланоспори мають дуже потовщену оболонку і червоне забарвлення. Статеве розмноження відбувається шляхом ізогамії, так само, як у стигеоклонію.
Нечисленні представники цього роду трапляються в Україні в різних водоймах, здебільшого навесні. Найбільш поширеним видом роду є ВгарагпаШіа %ІотегаІа - драпарнальдія грудкувата.
Родина трентеполіальні об'єднує виключно наземні гетеротрихальні водорості, таломи яких мають вигляд розгалужених ниток або одно- чи багатошарових паренхіматозних платівок. У трентеполіальних відсутні піреноїди, крохмаль синтезується у
незначній кількості і відкладається у вигляді дрібних гранул у стромі \ поропласту. Цитокінез відбувається за участю фрагмопласту, іумовлюючи утворення клітинної платівки зі справжніми порами та ііиіі імадесмами. Мітоз закритий.
Розмноження відбувається фрагментацією ниток, акінетами, чотириджгутиковими зооспорами та статевим шляхом. Життєвий цикл пшлодиплофазний, з ізоморфною зміною поколінь, причому зооспори утворює як гаметоспорофіт, так і спорофіт. Статевий процес і іогамний, здійснюється дводжгутиковими гаметами, які також здатні
роростати партеногенетично.
Трентеполіальні викликають червоне цвітіння кори дерев, паразитують на листках різноманітних деревних порід або є фікобіонтами лишайників. Біохімічною особливістю є здатність накопичувати велику кількість вторинних каротиноїдів (переважно - | ( каротину) та полігідроксиспиртів (поліолу та альдітолу). Типовий представник - ТгеМероИІіа.
Рід трентеполія - Trentepok.Ua. Таломи ТгепІероМіа мають иигляд коротких розгалужених сланких нигок, висхідні нитки редуковані. У багатьох видів кінцеві клітини помітно збільшені за
 розміром
і являють собою або зооспоракгії, або
гаметангії. Після приготування водних
препаратів можна спостерігати, як
протягом кількох хвилин з кінцевих
клітин виходять гамети або зооспори.
розміром
і являють собою або зооспоракгії, або
гаметангії. Після приготування водних
препаратів можна спостерігати, як
протягом кількох хвилин з кінцевих
клітин виходять гамети або зооспори.
Види роду часто зустрічаються на корі листяних порід дерев, викликаючи цегляно-червоне аеро- фітне "цвітіння".
Рис. 63. ТгеїЛероЬІіа.
А - клітини з гематохромом в центрі;
Б - зоосшрангій.
Порядок Едогоніальні - Ое<1оеопіаіе8
Об'єднує нитчасті та гетеротрихальні водорості, які за цитокінс- ііїм з участю клітинної платівки нагадують хетофоральні водорості, проте суттєво відрізняються від них наявністю особливого типу мітозу (едогонієвий мітоз), клітинним поділом з утворенням ковпачків, сте-
фаноконтними монадними клітинами (зооспорами, андроспорами та сперматозоїдами) та високорозвиненим оогамним статевим процесом.
Едогоніальні водорості можуть бути однодомними або дводомними. Життєві цикли у едогоніальних водоростей бувають двох типів: гаплофазні без зміни поколінь та гаплофазні з стадією мікроскопічного чоловічого гаметофіту - нанандрію. Види, які мають перший тип життєвого циклу, називають макрандрієвими, другий - нанандрієвими.
До порядку входить біля 500 видів водоростей трьох родів, з яких нижче розглядається один - Oedogonium .
Рід едогоніум - Осйо§општ. Таломи мають вигляд простих нерозгалужених ниток, що складаються з циліндричних або злегка головчастих клітин. Базальні клітини утворюють ризоїдоподібні вирости, якими водорість прикріплюється до субстрату. Верхівкова клітина тупа або загострена, зрідка витягнута в щетинку.
Клітинна оболонка гладенька або злегка хвиляста, і на окремих клітинах має характерні для даного порядку ковпачки. Утворення ковпачків є наслідком особливого мітозу та специфічного цитокшезу. На початку поділу біля верхівки клітини з'являється складка з дикгіо- сом, які, зливаючись, угворюють увігнуте в цитоплазму кільце. Далі ядро мітотично ділиться. Між дочірніми ядрами за способом фікоплас- ту починає формуватись клітинна перегородка, материнська оболонка у зоні кільця розривається. Кільце починає швидко розпрямлятися, за рахунок чого відбувається ріст верхньої нової клітини. Залишки оболонки материнської клітини лишаються на дочірній у вигляді ковпачка. Кількість ковпачків відповідає кількості поділів даної клітини.
При едогонієвому типі мітозу центріолі відсутні, центрами організації веретена виступають елементи ендоплазматичного ретику- люму. В анафазі дочірні ядра розходяться, а із залишків веретена утворюється фікопласт. У телофазі ядра помітно зближуються за рахунок скорочення залишків інтерзонального веретена. У фікопласті скупчуються пухирці, що відшнуровуються від ендоплазматичної сітки і містять матеріал майбутньої перегородки. Пухирці частково зливаються, і утворюється клітинна платівка з первинними порам.
Рис. 64. Оесі<^опіиіті.
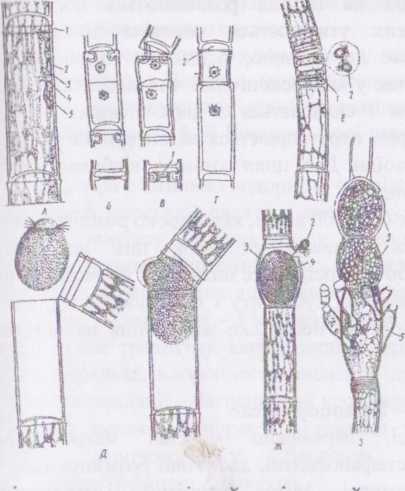 А
- Будова клітини:
А
- Будова клітини:
1 - ковпачок, 2 - стінка, З - хлоропласт (хроматофор), 4 - піреноїд,
5 - ядро.
Б,В,Г - поділ клітини з утворенням валика (1) і ковпачка (2):
Д - запліднення яйцеклітини (4) в оогонії (3) сперматозоїдом (2);
З - рослина з оогоніями (3) і чоловічими карликовими заростками - нанандріями (5).
У протопласті вегетативних клітин
розрізняється пристінним сітчастим хлоропласт з дрібними иіреноїдами, облямованими крохмальними платівками та одне ядро. Розмноження едогонію відбувається нестатевим або статевим шляхом.
Зооспори едогоніуму стефаноконтні. При утворенні зооспор одна з вегетативних клітин дещо здувається, клітинна оболонка ровривається; протопласт клітини виходить назовні і незабаром ниробляє фіброзне кільце з кількома десятками пар джгутиків, перетворюючись на стефаноконтну зооспору. Експериментальним шляхом встановлено, що на зооспори здатні перетворюватись будь-які вегетативні клітини, якщо отримати їх ізольовані протопласти.
Статеве розмноження у макрандрієвих та нанандрієвих видів иідбуваеться дещо різними шляхами.
У макрандрієвих видів як антеридії, так і оогонії розвиваються па звичайних вегетативних нитках. В антеридіях формується по два пісрматозощи, в оогонії - одна яйцеклітина. Сперматозоїди виходять з антеридію, підпливають до оогонію, через пору або тріщину в оболонні проникають до яйцеклітини та запліднюють її. Зигота одягається товстою оболонкою і переходить у стан спокою. По закінченні періоду і покою в зиготі відбувається редукційний поділ, з кожного гапло-
і іного ядра утвоюється зооспора, яка надалі проростає у нову нитку.
У нанандрієвих видів на нитках розвиваються оогонії та особливі спорангії, в яких утворюється чоловіча зооспора - андросиора. Вона підпливає до оогонію, осідає на нього або на сусідню клітину та проростає у мікроскопічний чоловічій гаметофіт, який називається нанадрієм і складається з двох-чотирьох клітин. Верхівкова клітина нанандрію перетворюється на антеридій, в якому розвиваються два сперматозоїди. Далі цикл розвитку відбувається так само, як і у макрандрієвих видів.
Oedogonium нараховує біля 400 видів, які широко розповсюджені у прісних водоймах. Як правило, розвиток цих водоростей починається у перифітоні або бентосі, проте нитки, які досягли значної довжини, можуть відриватися від субстраіу і продовжувати рости у вільному стані, утворюючи м'яке неслизьке жабуриння на поверхні води.
Клас Сифонофіцієві - Siphonophyceae
Об'єднує сифональні, переважно морські макроскопічні водорості. Зооспори часто стефаноконткі, джгутики розміщуються по спіралі; гамети двод*гутикові. Мітоз закритий. Цитокінез у вегетативних стадій відсутній, оскільки представники, мають неклітинну будову. У сифонофіцієвих виявлено специфічні ксантофіли
сифонеїн та сифоноксантин. Продуктом асиміляції замість крохмалю може бути полімер фруктози - полісахарид інулін.
У деяких сифонофіцієвих татом має два типи пластид, - забарвлені хлоропласти та безбарвні амілопласти. Амілопласти не приймають безпосередньої участі у фотосинтезі, а виконують функції накопичення запасних полісахаридів.
Розмноження здійснюється за допомогою дво-, або чотири джгутикових та стефаноконтних зооспор та статевим шляхом. При утворенні репродуктивних клітин fra спорангій або гаметангій перетворюється або весь талом, або його частина.
Порядок Бриопсидальиі - Brvopsidales
Об'єднує водорості, таломи яких не мають радіально- симетричної будови і являють собою одну велетенську клітину з багатьма (інколи - до кількох тисяч) ядрами. Центріолі зазвичай відсутні. Фотосинтетичний апарат представлений лише хлоропластами,
деяких видів карпогони розвивають коротку зачаткову трихогіну, яка має вигляд невеличкого сосочка.
Види цього роду поширені як у північних, так і в південних морях (зокрема, в Чорному морі), населяючи літораль та сублітораль. Живуть у прикріпленому стані на камінні, скелях, таломах великих морських водоростей. В Чорному морі порфіра вегетує з листопада по травень.
Порфіра є їстівною водорістю (це відображено у її народній назві
червоний морський салат), і введена у промислову аквакультуру. Зокрема, порфіру вирощують у Чорному морі на спеціальних морських фермах. Червоний морський салат вважається делікатесом, смак якого обумовлюють вільні амінокислоти та ізофлоризиди.
Клас Флоридові - ПогіїІеорИусеае включає водорості, у яких каргюгон має трихогіну, карпоспори утворюються різними шляхами у різних порядках, хлоропласти численні, дископодібні, без пірено'здів. Продукт асиміляції - багрянковий крохмаль (глікоген не утворюється). До класу входять водорості виключно з гетєротрихальним типом будови, причому у багатьох випадках утворюються псевдопаренхіматозні структури.
Рід батрахоснерм - ВаіїасИохрегтит. Всі види роду є прісноводними водоростями, що мешкають переважно у холодних
 гірських
річках або великих озерах. Таломи цих
водоростей можуть мати різні відтінки
як червоного, так і синьо-зеленого
забарвлення.
гірських
річках або великих озерах. Таломи цих
водоростей можуть мати різні відтінки
як червоного, так і синьо-зеленого
забарвлення.
Рис. 38. Батрахоспермум (Ва^асЬоїрегтит):
а - зовнішній вигляд талому; б — частина талому з карпоспорангіями
У ВсИгасЬохрегтит гаплоїдний макроскопічний гаметофіт розвивається на диплоїдному мікроскопічному спорофіті. Спорофіт у цієї водорості представлений однорядними розгалуженими нитками, у верхівкових клітинах яких може відбуватися мейоз (соматичний тип редукції). Після мейозу такі
клітини не перетворюються на тетраспорангії, а починають безпосередньо проростати у гаметофіти. Таломи гаметофітів оточені товстим шаром слизу, диференційовані на однорядну опірну нитку та мутовчасто розміщені бічні гілочки-асимілятори. Зовні гаметофіти виглядають як тендітні невеличкі (3- 8 см заввишки) синювато-зелені кущики, що складаються з окремих пагонів. Останні схожі на нитки з нанизаним на них намистом.
Сперматангії утворюються з апікальних клітин асиміляторних ниток, карпогони - на коротеньких карпогоніальних гілочках, які у різних видів розміщуються на різних частинах талому гаметофіту. Після запліднення зигота без періоду спокою ділиться і утворює щільну груп)' малоклітинних гонімобластів, у яких верхівкові клітини перетворюються на карноспори. Структура, утворена гонімобластами
з карпоспорами, має сферичну форму (вона трохи нагадує плід малини), зовні вкрита тонкою слизовою обгорткою і називається цистокарпієм. Карпоспори проростають в одноосьові розгалужені кущисті спорофіти.
Рід філофора — Phyllophora. З видів цього роду в Україні отримують агароїд, який називають "чорноморським агаром". Уздовж північно-західного узбережжя Чорного моря, між Одесою та Очаковим, знаходиться зона, де на глибині 5-60 м суцільні зарості утворює Phyllophora nervosa. Це найбільший у світі філофорний фітоценоз, який називають філофорним полем Зернова.
Таломи Phyllophora кущисті, представлені сланкими "пагонами", від яких підіймаються вертикальні стебельця. Верхня частина кожного стебельця сплощена, розростається по краям, і утворює пластину з потовщеною середньою частиною - ребром. Пластина рясно галузиться, часто по краям та на ребрі несе дочірні, молодші пластини. Гетра-
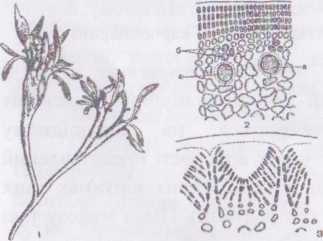 споранпі
та статеві органи розвиваються у
нематеціях, розташованих на поверхні
пластин вздовж ребра.
споранпі
та статеві органи розвиваються у
нематеціях, розташованих на поверхні
пластин вздовж ребра.
Рис. 39. Будова і розмноження філофори РЬуїіорЬога:
- рослина з нематеціями;
- поперечний розріз талому ;
- сперматангії на поперечному зрізі.
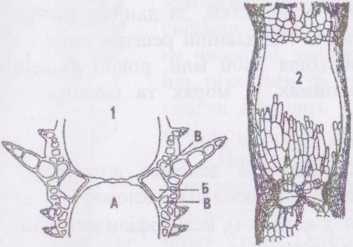
Рід цераміум - Сегатіипь Види роду мають вигляд невеличких кущиків, утворених дихотомічно розгалуженими нитками із загнутими всередину вильчаетими верхівками. 'Галом утворений однорядною опірною ниткою, у якої зона контакту суміжних клітин вкрита корою. У деяких видів кора вкриває весь талом, проте частіше середня частина клітин опірних ниток лишається відкритою. Як наслідок, рослина набуває членистого вигляду - прозорі відкриті ділянки опірних клітин чергуються з ділянками, вкритими корою, що забарвлена в інтенсивний червоний колір.
Кора являє собою асиміляторні короткі нитки, що щільно притискаються до оболонок клітин опірної нитки. Деякі асиміляторні нитки видозмінюються у щетинки або шипи. Сперматангії та карпогоніальні нитки у гаметофітів та тетраспорангії у спорофітів розвиваються з клітин кори.
 Сегатіит
зустрічається в усіх широтах, але масово
розвивається переважно у теплих морях,
зокрема в Чорному та Азовському.
Сегатіит
зустрічається в усіх широтах, але масово
розвивається переважно у теплих морях,
зокрема в Чорному та Азовському.
Рис. 40.
Будова Сегатіит:
- поздовжній зріз через вузол:
а) центральна клітина;
б) перицентральна клітина;
в) вторинні корові клітини;
- частина слані.
Рід полісифонія - Роїувіріїопіа. Таломи цієї водорості також кущисті і мають членисту будову, але іншого типу, ніж у Сегатіит. Нитку утворюють два типи клітин - центральні, що виконують опірну функцію, та перицентральні, які здійснюють фотосинтез і утворюють репродуктивні органи: тетраспорангії у спорофітів, сперматангії та карпогоніальні гілки у гаметофітів.
Перицентральні клітини розташовуються правильними ярусами, причому один ярус перицентральних клітин оточує однорядним кільцем одну центральну клітину. Перицентральні та центральну клітини одного ярусу називають, відповідно, периферичними та центральним сифонами.
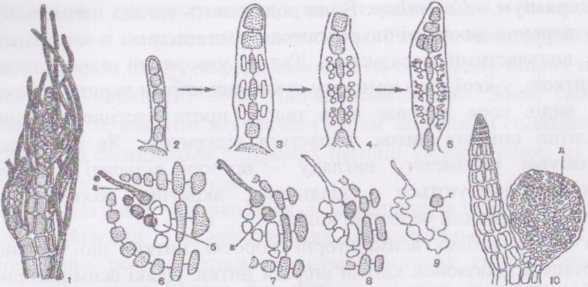
Рис.
41.
Будова і розмноження Polysiphonia:
1
-
вершина гілочки; 2-5
-
схема розвитку сперматангіїв на
трихобласті; 6-10
-
схема розвитку гонімобласта; а - карпогон,
б - несуча клітина, в - ауксилярна клітина,
г - перша клітина гонімобласта, д —
цистокарп.