

Відділ Діатомові водорості - ВасШагіорІїуіа
Відділ налічує більше 20 тис. видів. Діатомові водорості виникли порівняно недавно - біля 130 млн. років тому, у крейдяний період фанерозою. У теперішній час діатомові перебувають у стані біологічного прогресу і є домінуючою групою у морських та прісноводних біотопах. Всі діатомові водорості - це кокоїдні одноклітинні або колоніальні організми, розміри яких коливаються в діапазоні 4-2000 мкм при середніх значеннях 20-50 мкм.
Фотосинтетичні пігменти представлені хлорофілами а та с, Р- та є-каротинами. З ксантофілів переважає жовтий пігмент фукоксантин, який обумовлює коричневе забарвлення пластид.Основним продуктом асиміляції є хризоламінарин, додатковими - олія та волютин. Продукти асиміляції відкладаються або безпосередньо у цитоплазмі, або у вакуолях.
Клітинні покриви представлені кремнеземовим панциром, що розташовується назовні від плазмалеми. Панцир у всіх діатомових водоростей утворений двома половинами: більшою - епітекою, та меншою - гіпотекою. Кожна половина панциру складається зі стулки та пояскового обідка, завдяки цьому панцир має вигляд коробки (гіпотеки), яка накрита кришкою (епітекою). Зона перекривання пояс- кових обідків епітеки та гіпотеки називається пояском. Панцир має досить складну структуру, специфічну для кожного роду та виду діатомових водоростей. Форма панциру у діатомових водоростей досить різноманітна, і характеризується, в першу чергу, типом симетрії стулки: радіально симетричні та двобічносиметричні, а також типом розташування ареол, швів і різних шипиків, виростів тощо.
До внутрішнього боку панцира щільно притиснута плазмалема.
Чимало видів діатомових водоростей здатні виділяти слиз. За допомогою слизу водорості утворюють слизові грубки, різноманітні слизові ніжки, з'єднуються у колонії. У водоростей, які мають шов, слиз відіграє надзвичайну роль у процесі активного ковзаючого руху клітини.
Ядро одне, розташовується в центрі клітини. Біля поверхні ядра, як правило, розташовується 1-2 комплекси Гольджі. Мітоз у діатомових водоростей відкритий, центріолі відсутні. Замість них функції центру організації мікротрубочок виконують особливі полярні диски.
Фотосинтетичний апарат представлений вторинно симбіотичними пластидами родофітного типу.Тилакоїди по три зібрані у ламели; безпосередньо під оболонкою хлоропласту розташовується оперезуюча ламела. Досить часто в хлоропласті є голий піреноїд. Мітохондрій зазвичай кілька. Вони розміщуються у периферичній частиш’ клітин і мають лише трубчасті кристи.
Вакуолярний апарат включає 4 типи вакуолей: з клітинним соком, волютином, хризоламінарином та олією.
Монади і стадії представлені виключно голими одно джгутиковими сперматозоїдами, і відомі лише у кількох родів діатомових водоростей.
Діатомові водорості мають виключно кокощний тип морфологічної структури тіла, хоча в деяких випадках ниткоподібні колонії діатомей можуть бути помилково ідентифіковані як нитчасті форми.
Представники відділу розмножуються вегетативно (поділом клітин надвоє) та статевим шляхом. Всі діатомеї мають єдиний тип життєвого циклу - диплофазний цикл з гаметичною редукцією, без зміни поколінь та зі специфічною ростовою стадією - ауксоспорою.
Внаслідок вегетативного поділу надвоє кожна дочірня клітина отримує половину панцира, але добудовує кожна з них тільки гіпоте- ку. Клітина, що отримала у спадок материнську епітеку, зберігає розміри материнської клітини. У другої клітини половина панциру, що у материнської клітини була гіпотекою, стає епітекою. Оскільки кремнезем не здатний розтягуватись, то в принципі розміри клітин у популяції мали б швидко зменшуватись. Проте, в дійсності такого не відбувається, оскільки у діатомових водоростей існують певні механізми підтримання сталих розмірів. Серед них до провідних належать механізми відтворення оптимальних розмірів через стадію ауксоспори, переважно внаслідок статевого процесу. Ауксоспора (або спора, що росте) - це особлива клітина, яка здатна до росту, і най частіше вони виникають після статевого процесу з зиготи, що проростає.
При статевому процесі дві клітини вкриваються спільним слизом, протопласти збільшуються у розмірах і в результаті редукційного поділу формують гамети. В залежності від типу статевого процесу, характерного для кожного виду, життєздатними лишаються одна, дві, або чотири гамети. У водоростей з актиноморфними стулками найчастіше зустрічається оогамний статевий процес. У водоростей з зигоморфними стулками найбільш поширеними типами статевого процесу є ізо- та гетерогамія.
Особливим способом статевого процесу у діатомей є автогамія. У такому процесі бере участь лише одна клітина. Її ядро редукційно ділиться, далі два гаплоїдних ядра дегенерують, а два інших зливаються, відновлюючи диплоїдне ядро і утворюючи зиготу, що перетворюється на ауксоспору.
Спочиваючі стадії представлені двостулковими кремнеземовими цистами.
Поділ на класи проводиться, в першу чергу, за типом симетрії стулок (актиноморфні чи зигоморфні) та наявністю шва. З цим комплексом ознак корелює чимало ознак, пов'язаних з будовою протопласту та особливостями статевого процесу. Відділ поділяють на два класи - Центричні (СегіїгорЬусеае) та Пенатні (РсппаЮрИусеае)
Клас Центричні - Сепй-орЬусеае об'єднує водорості з актиноморфними стулками, які завжди позбавлені шва. Лише у центричних діатомей виявлено оогамні статеві процеси та рухливі сперматозоїди. Представники цього класу мешкають переважно у планктоні морів та прісних водойм. Типовий представник - Меіоша.
Рід мелозіра - Меіохіга Водорість надзвичайно характерна для планктонних угруповань прісних водойм. Колонії мають вигляд нероз- галужених ниток, які складаються з циліндричних клітин, з'єднаних між собою за допомогою слизових валиків, шипів або зубців. У Меіоиіга досить детально досліджено статевий процес, який представлений оогамією. На початку статевого процесу в одній з клітин відбувається мейоз. При цьому після першого поділу (мейоз 1) одне з ядер дегенерує, і не переходить у фазу мейозу 2. Друге, життєздатне ядро, ділиться, і знов одне з ядер дегенерує. При поділах у фазах мейоз
І та 2 цитокінез не відбувається. Таким чином, у жіночому гаметангії утворюється одна яйцеклітина.
В клітині, що виконує функцію чоловічого гаметангію, протопласт набрякає, обумовлюючи скидання панциру. Далі на поверхні протопласту з’являються чотири джгутики, відбувається мейоз і утворюється чотириядерна гола сперматогенна клітина, що зберігає всі чотири джгутики Далі одночасно від сперматогенної клітини відбруньковуються чотири одноджгутикові одноядерні сперматозоїди. Брунькування відбувається таким чином, що в гамети не потрапляють пластиди: вони залишаються в центрі сперматогенної клітини, утворюючи т.зв. залишкову клітину, що позбавлена ядра і незабаром руйнується. Далі сперматозоїди запліднюють яйцеклітину, яка перетворюється на ауксоспору.
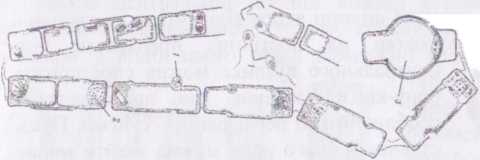 Рис.
ЗО.
Melosira.
Рис.
ЗО.
Melosira.
- розвиток сперматозоїдів;
- розвиток яйцеклітини; 3 - утворення ауксоспори.
Види роду мело- зіра широко представлені в планктоні ставків, озер, річок, заплавних водойм. Серед представників роду є також і літорально-бентосні організми, деякі види здатні розвиватись на зрошу ваних скелях.
Клас Пенатні - Pennatophyceae об'єднує водорості з зигоморфними стулками, які позбавлені швів (Araphales), або мають шви (Monoraphales, Diraphales).
 Рід
фрагілярія -
Fragilaria
Клітини водорості здебільшого
утворюють колонії у вигляді стрічок, в
яких панцири з'єднуються стулками. При
вигляді з пояска окремий панцир має
видовжено- прямокутну форму; зі стулки
панцир видовжений, білатерально-симетричний.
Види роду є L переважно
мешканцями перифітонних та щ
бентосних угруповань прісних водойм.
Рід
фрагілярія -
Fragilaria
Клітини водорості здебільшого
утворюють колонії у вигляді стрічок, в
яких панцири з'єднуються стулками. При
вигляді з пояска окремий панцир має
видовжено- прямокутну форму; зі стулки
панцир видовжений, білатерально-симетричний.
Види роду є L переважно
мешканцями перифітонних та щ
бентосних угруповань прісних водойм.
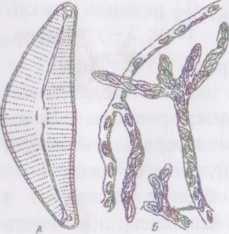
s Рис. 31. Fragilaria crotonensis.
А - колонія; Б - панцир зі стулки
Рід астеріонела - Asterionella. За будовою , U панциру вельми схожа з фрагілярією прісноводна
планктонна водорість АяГегіопеПа, яка, проте, добре відрізняється за будовою колоній. В них клітини з'єднані у зірчасті колонії за допомогою слизу та дрібних шипіків, розташованих на полюсі стулки.
 Рис.
32. АвіегіопеНа.
Рис.
32. АвіегіопеНа.
А - панцир зі стулки;
Б - загальний вигляд колонії
/
 центрального
вузлика можна спостерігати одну-кілька
крапок, що представляють особливий тип
перфорацій - стигми. Представників
цього роду можна знайти майже в будь-якій
прісній водоймі, переважно в обростаннях
вищих водних рослин.
центрального
вузлика можна спостерігати одну-кілька
крапок, що представляють особливий тип
перфорацій - стигми. Представників
цього роду можна знайти майже в будь-якій
прісній водоймі, переважно в обростаннях
вищих водних рослин.
Рис. 33. СутЬеІІа:
А - панцир з боку стулки;
Б — колонія в слизистій трубці
 Рід
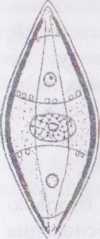
навікула - Ламсм/а Види одного з
найчисленніших родів порядку - Nауісиїа
- мають досить просту будову: стулки за
обрисами ланцетоподібні, і нагадують
човник. По середині осьового поля
проходить прямий щілиноподібний шов.
На поверхні стулок розташовуються
паралельні, радіальні або конвергентні
штрихи, утворені правильними рядами
ареол. Центральне поле відносно невелике,
і не доходить до країв стулки. При вигляді
з пояска клітини видовжено- прямокутні.
Пояскові обідки прості, і не мають
вставних обідків. Хлоропластів два,
вони пластинчасті, розташовуються
вздовж пояскових обідків на різних
боках стулки і містять по одному
паличкоподібному піреноїду.
Рід
навікула - Ламсм/а Види одного з
найчисленніших родів порядку - Nауісиїа
- мають досить просту будову: стулки за
обрисами ланцетоподібні, і нагадують
човник. По середині осьового поля
проходить прямий щілиноподібний шов.
На поверхні стулок розташовуються
паралельні, радіальні або конвергентні
штрихи, утворені правильними рядами
ареол. Центральне поле відносно невелике,
і не доходить до країв стулки. При вигляді
з пояска клітини видовжено- прямокутні.
Пояскові обідки прості, і не мають
вставних обідків. Хлоропластів два,
вони пластинчасті, розташовуються
вздовж пояскових обідків на різних
боках стулки і містять по одному
паличкоподібному піреноїду.
Рис. 34. ^уісиїа. Вигляд талому з боку стулки.
Рід нінулярія - Ріппиіагіа. Стулки видів роду Ріппиіагіа мають форму, наближену до видовжено-еліпсоїдної. Шов у піннулярії дещо хвилястий. З внутрішнього боку стулки панцир має систему грубих потовщень - ребер, які дещо нагадують поверхню пральногдошки, і в оптичний мікроскоп можуть бути помилково прийняті за штрихи. Між ребрами панцир перфорований рядами дрібних ареол. При вигляді з пояска панцир правильно-прямокутний, причому в цій позиції добре помітно, що ребра є саме системою внутрішніх потовщень стулки, а не її перфорацій.
Хлоропластів два, стрічковидні, розташовуються вздовж поясків з протилежних боків стулки і мають дуже порізані краї. У деяких видів протилежні стрічки хлоропласту під гіповальвою з'єднуються невеликим містком, завдяки чому хлоропласт набуває Н-подібної форми. В центральній частині кожної стрічки у багатьох видів розташовується по одному голому еліпсоїдному піреноїду. Оскільки панцири у видів цього роду переважно великі, клітини не здатні
 утримуватись
у товщі води, через що мешкають або у
бентосних, або у перифітон- них угрупованнях
прісних водойм.
утримуватись
у товщі води, через що мешкають або у
бентосних, або у перифітон- них угрупованнях
прісних водойм.
Рис. 35. Ріппиіагіа.
А - панцир з боку стулки Б - панцир з боку пояска;
В - клітина з боку стулки Г - клітина з боку пояска Д - поділ пінулярії: дві дочірні клітини з боку пояска