

1. Пролог к прологу. До человека.
1. 0. Предварительные замечания.
Конечно, никакой семиотики в те древнейшие времена, к которым я сейчас собираюсь обратиться, не было, от тех времен до нас дошли только продукты знаковой деятельности. Мне кажется, однако, что без исторической перспективы события видятся более плоскими, а, следовательно, искажаются. В этом параграфе в центре моего внимания будут стоять проблемы овладения открытыми знаковыми системами, рефлексии и осмысления знаковой деятельности, первые стадии знаковой рефлексии, связи знаковой рефлексии с другими ее типами.
1. 1. Проблемы глоттогенеза в научной литературе. Методика исследования.
Глоттогенез и генезис человеческой культуры являются центральными проблемами антропогенеза, поскольку человеческий естественный язык все специалисты по антропогенезу называют одной из главных характеристик, отличающих человека от животного, а человеческая культура в большой своей части является чем-то производным от естественного языка. Проблема описания процесса возникновения языка настолько сложна, по ней было написано такое количество спекулятивных работ, ничего общего не имеющих с научным описанием, что выведенное из себя Парижское лингвистическое общество даже ввело мораторий на принятие к рассмотрению работ, посвященных этой теме. Действительно, до недавнего времени в общем не существовало фундированной научной методики, которая позволяла бы получать сколь-нибудь правдоподобные результаты в этой области. Рассуждения о происхождении языка базировались, в основном, на более или менее остроумных гипотезах авторитетных философов. Философия же, даже самая современная, несет на себе печать религиозных представлений, которые в какой-то момент развития религиозного мифа, в той его части, которая касается происхождения человека, в их порыве вверх, к божественному, навсегда отделили человека от остальной природы, от животного и растительного мира. Голос Дарвина до сих пор слаб в этой области знания, как впрочем и во всех других областях гуманитарного знания - в истории, социологии, лингвистике, умозрительной психологии, искусствоведении и пр.
Все эти теории можно было бы подразделить на две большие части: теории, по которым язык был дан человеку богом, и теории, по которым человек обрел язык благодаря самому себе. Так, согласно Ригведе речь была сотворена богом священной речи Брихаспати совместно с семью певцами – риши, в Библии специально вопрос о происхождении речи не обсуждается, говорится лишь, что Бог создал землю и все, что есть на ней, произнося имена тех объектов, которые создавались, говорится также о том, что имена растениям и животным дал первочеловек Адам.
У древнегреческих философов суждения о происхождении речи вытекали из их общефилософских взглядов. Имеется четыре основные точки зрения на происхождение языка: теория, по которой имена вещам были даны так называемыми номотетами, теория звукоподражания, теория междометий и теория социального договора. Согласно первой точке зрения, лучше всего представленной в диалоге Платона Кратил, имена вещам дал закон, а закон создает законодатель (номотет): «Сократ. Ты не знаешь, кто передал нам имена, которыми мы пользуемся? Гермоген. Право не знаю. С. Не кажется ли тебе, что их дал закон? Г. Похоже, что так. С. В таком случае учитель, когда пользуется именем, пользуется трудом законодателя? Г. Я полагаю» (Платон. Кратил. 188д-е). Однако далее у Платона есть к этим утверждениям еще несколько существенных уточнений. Оказывается за мастером имен должен присматривать тот, кто этими именами пользуется, т. е. диалектик. Однако, еще дальше мы узнаем, что имена бывают двух типов: те, которые вещам дали люди, и те, которые вещам дали боги. Правильнее – те, которые вещам дали боги.
Согласно второй точке зрения, идущей от стоиков, имена вещам дали люди, с течением времени имена менялись, но восходят они все к звукоподражаниям.
Третью точку зрения развивали противники стоиков – эпикурейцы. По их воззрениям все имена произошли из междометий. Древние люди, по их мнению, превратили в естественные звуки инстинктивные животные вопли, которые затем и стали именами вещей.
Четвертую точку зрения развивал Демокрит, который полагал, что имена вещам люди дали по взаимному соглашению, договору. В изложении последнего античного неоплатоника – Прокла его концепция выглядит следующим образом: «Демокрит… говорил, что имена существуют по установлению, и доказывал это, приводя четыре довода: 1) одноименность – гомонимия, а именно то обстоятельство, что различные вещи обозначаются одним и тем же названием; 2) многоименность – полионимия: различные названия применяются к одному и тому же предмету, заменяя друг друга, а это невозможно, если названия вещей существуют по природе; 3) переименования вещей: ведь если бы имена были от природы, то почему бы Аристокла переименовали в Платона, а Тиртама в Феофраста; 4) отсутствие соответствия в словообразовании: например, от слова «мысль» можно образовать глагол «мыслить», почему же от слова «справедливость» нельзя образовать глагол «справедливить». Значит имена действительно возникли случайно, а не присущи вещам по природе» (цит. по Перельмутер 1980, стр.119).
В XIX в. появилась теория Нуаре о происхождении языка из выкриков, сопровождавших коллективный труд. Сходной точки зрения придерживался и Ф. Энгельс.
Как, на мой взгляд, правильно замечает Н. Н. Воронцов, «Я бы не стал здесь включать в число человеческих признаков труд, как то делал Ф. Энгельс в своей известной книге, поскольку отличить охотничий и собирательский труд древнего человека от охотничьей и собирательской активности диких млекопитающих (не только приматов) совсем не просто [Сноска: Орудийная деятельность свойственна ряду животных, не только приматам. Известно, что представители одного из видов вьюрков с помощью отломленных от кактусов колючек зондируют кору деревьев, добывая этим орудием личинок насекомых. Калан (Enhidra lutris) использует небольшие камни для разбивания панцирей морских ежей. Ныряя за очередным ежом, калан нередко сохраняет свое орудие подмышкой, с тем, чтобы, вынырнув с очередной жертвой, вновь использовать камень.]» (Воронцов 1999, стр. 16). Если же (тяжелый) труд человека перестает противопоставляться активности диких животных, становится сразу непонятным, почему самое трудолюбивое на свете существо - пчела - не выучилась производить трудовые выкрики для координации действий своих собратьев (улей живет по строжайшему распорядку, кроме того, у пчел, в отличие от людей, видимо, нет понятия отдыха, они все время в действии, как и муравьи, и другие общественные насекомые).
В последнее время в Западной Европе появился целый ряд новых концепций происхождения языка, связанных с эволюционной теорией Дарвина. Наиболее авторитетные из них уходят своими корнями в идеи, высказанные в свое время Э. Кассирером в его «Философии символических форм» и «Опыте о человеке» (см. Кассирер 1998). Уже в беглом обзоре эволюции взглядов европейских философов на природу человека Кассирер практически предсказал сильное влияние естественно-научных достижений на гуманитарные исследования, в центр которых он ставил развитие символических форм. В истории развития человеческого самопознания Кассирер выделил 4 основных этапа: первый этап открывается Сократом и стоиками, поставившими человека, человеческий разум в центр мироздания, второй этап был христианско-неоплатонической доктриной Блаженного Августина, который отнес величие человеческого разума к эпохе, предшествующей грехопадению наших прародителей – Адама и Евы, а современному человеку оставил только надежду на то, что божественный разум может к нему вернуться только через божественную благодать, третий этап начинается с открытия Коперником гелиоцентрической системы, после которого претензия «человека на то, чтобы быть центром вселенной потеряла основания. Человек помещен в бесконечном пространстве, в котором его бытие кажется одинокой и исчезающе малой точкой. Он окружен немой Вселенной, миром, который безмолвно безразличен к его религиозным чувствам и глубочайшим моральным запросам» (Кассирер 1998, стр. 457 – 458); первым постулатом нового духа времени «было устранение всех искусственных барьеров, которыми человеческий мир прежде был отделен от остальной природы» (Кассирер 1998, стр. 457). Четвертый этап по Кассиреру начинается с Дарвина: «В первой половине XIX в. работали еще такие метафизики, как Гербарт, и такие психологи, как Г. Т. Фехтнер, которые лелеяли надежду основать математическую психологию. Но эти проекты быстро исчезли после появления книги Дарвина «Происхождение видов». С этих пор подлинная сущность антропологической философии определилась раз и навсегда. После неисчислимых бесплодных попыток построения философии человека она обрела, наконец, твердое основание. Мы не чувствуем больше потребности строить воздушные замки, предаваться спекуляции, ибо мы вообще теперь не стремимся дать общее определение природы или сущности человека. Наша задача заключается в сборе эмпирических данных, которые щедро предоставляет в наше распоряжение общая теория эволюции» (Кассирер 1998, стр. 462).
Кассирер понимал, что центральным пунктом в теории глоттогенеза должен стать, с одной стороны вопрос о наследовании, об истоках развития языка, с другой стороны, - вопрос о скачке, о мутации, которая привела к преобразованию дочеловеческих семиотических систем в человеческий язык.
Осознание того, что проблему происхождения языка следует увязать с антропогенезом, с эволюцией в целом привел в настоящее время к развитию нескольких дисциплин, объектом изучения которых являются а) самые ранние этапы развития знаковых систем, а эти этапы в связи с осознанием семиотической природы генетического кода увязываются сейчас с этапами возникновения жизни на Земле; б) семиотические системы низших организмов (знаковые системы одноклеточных; знаковые системы растений – фитосемиотика; знаковые системы грибов – микосемиотика; знаковые системы животных – зоосемиотика); в) семиотические системы клеточных образований организма (генетический код, как семиотическая система; знаковые процессы имунной системы; семиотика нейронов, семиотические процессы, происходящие в нервной системе); эволюция знаковых систем ( в том числе и языка).
Мое исследование примыкает к той группе глоттогенетических штудий, которые стараются опираться на факты всех наук, которые способны пролить свет на тайну происхождения языка и которые признают следующие исходные постулаты: 1) язык человека представляет собой функционально выделенный психо-физиологический механизм, предназначенный для построения языковых текстов, речевых произведений; речевое произведение всякий раз является компонентом речевого поведения, а речевое поведение является компонентом поведенческой программы, ставящей себе задачу достигнуть некоторой поведенческой цели; 2) язык развивался параллельно с развитием нервной системы и поведенческих программ, которые он обслуживал; 3) естественный звуковой язык человека - результат эволюции семиотических систем; он развился из старших семиотических систем; его формирование зависело от формирования физических, физиологических и психологических подсистем; 4) назначение языка - обслуживать социальную жизнь языкового коллектива, следовательно усложнение языка идет параллельно усложнению социума, более того процессы развития языка и развития социума взамообусловлены.
Отрыв глоттогенетической теории от эволюционного учения, отрыв интерпретации языковых фактов от этологических, нейрофизиологических, физиологических и др. факторов ведет к фантастичности многих теорий глоттогенеза и тому большому разнобою, который, например, характеризует гипотетические утверждения по поводу таких важных вопросов, как датировка преобразования семиотической системы гоминид, в общем, мало чем отличавшейся от семиотических систем, которыми пользуются животные других видов, в естественный человеческий звуковой язык. Одни исследователи соотносят этот процесс с так называемым Homo habilis (≈2 млн. лет до р. Х.), другие соотносят его с определенным периодом существования Homo erectus (≈500 000 лет до р. Х.), третьи - только с Homo sapiens sapiens (≈50 000 лет до р. Х.). Одни признают, что процесс этот должен был быть монотонным и непрерывным, другие считают, что при переходе от семиотической системы, аналогичной зоосемиотическим, к языку был скачок. Одни считают, что звуковому языку предшествовал в филогенезе жестовый язык, другие так не считают, и т. д.
Несомненно, глоттогенез является комплексной проблемой, решить которую можно только используя достижения различных наук: археологии, палеоантропологии, палеоневрологии, нейрофизиологии, нейроанатомии, психологии, лингвистики, этологии и др. Мне кажется, что позитивный сдвиг в работах, посвященных этой теме, произошел тогда, когда появились комплексные исследования, оперирующие данными всех или почти всех наук, которые имеют отношение к данной проблематике. Одной из первых подобного рода работ, содержавших сводку наиболее интересных и перспективных результатов, которые были получены в рамках наук, перечисленных выше, явилась работа Иванов 1976. Вяч. Вс. Иванов предложил одновременно и методы исследований, которые и на мой взгляд должны привести к наиболее правдоподобным результатам. Я имею в виду, во-первых, последовательно проведенный метод компаративных исследований, в которых бы сравнивались данные анатомии, нервной организации, этологии, семиотических систем и пр., с одной стороны приматов, с другой стороны, ископаемых гоминид, с третьей стороны человека. Во-вторых, последовательно же проведенный метод внутренней реконструкции, предполагающий внимательное исследование нестандартных элементов, возникающих в механизмах поведения, коммуникации и пр. На мой взгляд, эти методы позволяют гораздо более строго отбирать в качестве наиболее правдоподобных гипотезы, касающиеся глоттогенеза. Покажу это на примере нескольких известных в науке предположений.
Если, скажем, говорить о времени возникновения языка, то в первую очередь следует договориться о том, чем отличается естественный человеческий звуковой язык от языка животных, в том числе и от семиотических систем, которыми пользуются приматы. Только тогда обнаруженные с помощью изощренных изыскательских методов признаки владения ископаемым человеком некоторой семиотической системой можно оценить с точки зрения ее отнесенности к классу естественных языков и только тогда, когда мы убедимся, что семиотическая система, обнаруженная с помощью этих методов, относится к естественным языкам, мы можем обратиться к вопросу о том, к какому времени она относится, чтобы затем выдвинуть гипотезу о том, что возникновение языка у человека относится к такому-то периоду истории.
Пожалуй, сейчас наука располагает (если вынести данные археологии за скобки) восемью более или менее обнадеживающими источниками сведений о типе семиотических систем, которыми владели ископаемые люди: 1) реконструкция гортани ископаемого человека, которая стала возможной благодаря работам палеозоологов и палеоантропологов; 2) реконструкция структур мозга ископаемого человека, которая стала возможной благодаря исследованиям по палеоневрологии; 3) реконструкция структуры органов слуха ископаемого человека; 4) реконструкция позвоночного канала, в котором располагается спинной мозг, позволяющая судить о том, каковы были возможности спинного мозга в управлении дыхательными операциями (речевой режим дыхания управляется довольно сложными операциями по модулированию подачи воздуха к голосовым связкам, для этого требуется достаточно сложная система управления легкими, что сказывается на диаметре позвоночного канала и конфигурации позвонков); 5) исследования, относящиеся к появлению и развитию речи у ребенка (развитие языка в онтогенезе); факты, которые предоставляет в наше распоряжение эта область знания могут пролить свет на проблему происхождения языка, если учесть параллель, существующую между теми связями, которые существуют между эволюцией живых существ и процессом развития зародыша, а также между развитием языка в филогенезе и доречевой стадией развития ребенка; 6) результаты исследований, полученных в рамках сравнительно-исторического языкознания; 7) результаты исследований древнейших и «рудиментарных» компонентов естественного языка, в частности, - идеофонов, или звукоподражаний; 8) данные мифологии.
Сведения, полученные из каждого из этих источников уже позволяют как-то ограничить спекулятивные рассуждения на интересующую нас тему и ввести их в некоторые научные рамки. Так, сравнительный анализ гортани позволяет установить, что у взрослого человека в отличие от обезьяны (и от ребенка в возрасте до года) надгортанник располагается довольно низко. Благодаря этому образуется мощный, хорошо управляемый резонатор, позволяющий человеку издавать довольно громкие и разнообразные звуки. Надгортанник осуществляет очень важную речевую работу: он модулирует звуковой сигнал таким образом, чтобы громкость гласных не заглушала громкость согласных. Из этого следует, что высокое положение надгортанника не позволяет осуществлять речепроизводство. Следовательно, если реконструкция строения гортани покажет, что, скажем, у австралопитеков надгортанник расположен высоко, как у обезьяны, мы можем смело говорить, что звуковой речи у них не было. При этом положительных заключений на основании низкого положения надгортанника мы делать не можем.
Ссылки на то, что гортань австралопитеков или Homo habilis, или Homo erectus была способна воспроизводить такие-то, или такие-то гласные или согласные не могут ни в какой степени подтвердить или опровергнуть гипотезу о том, что эти ископаемые люди владели языком. Обращение, например, к моделирующим устройствам, позволяющим понять, что может произносить гортань австралопитека и чего она произносить не может, ничего не дает. Ведь в человеческом языке главное не звуки, а модели звуков - фонемы. Об этом писали еще основатели семиотики Ч. С. Пирс и Ф. де Соссюр. Одна и та же модель звука - фонема - может иметь бесчисленное количество звуковых репрезентаций. Так, например, фонема /а/ в русском языке может быть произнесена миллионом различных тембров, на определенном, но континуальном, участке тонового спектра, т. е. более высоким и более низким по высоте тона голосом, в различных режимах работы голосовых связок, начиная от шепотной речи, кончая визгом, кроме того, еще и артикуляционные характеристики этой фонемы могут варьироваться в зависимости от позиции в фонетическом слове, в зависимости от окружения, в котором она встречается. Так, в словоформе мама [m o´ão m o˜∂] в первой позиции эта фонема репрезентируется долгим ударным, назализованным, огубленным /а/, во второй позиции - редуцированным кратким, назализованным, огубленным шва. В словоформе такой [tâkoóoî] она репрезентируется закрытым неогубленным, неназализованным /а/. Для других языков эти и подобные им различия могут означать, что соответствующие звуки относятся к разным фонемам. Из этих замечаний следует, что главное для звука, воспроизводящего фонологическую модель, принадлежащую семиотической системе, имеющей статус естественного языка, вовсе не то, какая у него экспонента, а то, к какому классу экспонент он относится, а класс экспонент определяется фонемой, задающей класс эквивалентности на множестве звуков, произносимых членами данного языкового сообщества. Один и тот же звук в естественном языке может относиться к разным классам эквивалентности. Так одно и то же |â| в такой и покой [p оâ kо ó оî] относятся к репрезентациям разных фонем - /а/ и /о/.
Кроме сказанного выше, важно еще и то, какие параметры в звуке считаются существенными, или, как говорят лингвисты, релевантными. Скажем, в крике грудного ребенка, изображаемого на письме в виде а-а-а-а, важно не то, что это не э-э-э-э-э, а то, в каком режиме работают его голосовые связки, важен сам тембр: если ребенок кричит «противным, вредным» голосом, который невозможно долго выдержать, чтобы не подойти и узнать, что с ним произошло, то это значит, что ребенок чем-то недоволен, если в его крике нет напряжения, то это может значить, что он просто тренирует голосовые связки и не требует вашего вмешательства в обстоятельства, определяющие его поведение. В этом случае главными характеристиками звука являются составляющие тембра. Если же звук используется как репрезентанта фонемы, то важными оказываются именно артикуляционные характеристики, причем не все, а совершенно определенные, существенные для смыслоразличения в данном языке.
Определить по экспоненте может ли она быть репрезентацией фонемы, или нет, по археологическим данным нельзя. Моделирование здесь не поможет. Даже если бы мы точно узнали, что ископаемые люди столь раннего периода могли произносить так называемый членораздельный звук. Это тоже бы нам ничего не дало для подтверждения гипотезы о существовании у ископаемого человека данного периода естественного языка или для ее опровержения. Один членораздельный звук ничего не может сказать о комбинации звуков, а комбинируемость - главный параметр, отличающий язык человека от семиотических систем животных.
Важным критерием является и то, образуют ли звуки в комбинации звуковую цепочку или нет. Звуки в семиотических системах животных не комбинируются в звуковые цепочки, не квантуются слоговым механизмом и, таким образом, не образуют слогов, не образуют морфов, не образуют слов. Они, как правило, равны целому сообщению. Вообще наличие или отсутствие уровневой организации в тексте и есть главный критерий, отличающий человеческую речь от семиотических текстов животных3. Если этого не учесть, бессмысленно говорить о том, когда у гоминид появилась звуковая человеческая речь.
Далее. Физиологическими, в частности, нейрофизиологическими условиями, без которых нельзя говорить о возможности речепроизводства в указанном выше смысле, являются наличие видимой физической асимметрии полушарий головного мозга, которые свидетельствуют о развитых зонах (в мозгу), которые отвечают за понимание и синтез речи, я имею в виду зону Брока и зону Вернике4. Зоны Вернике и Брока в свою очередь не могут появиться в человеческом мозгу без предварительного развития доминантности правой руки, без появления у человека центров координирующих действия правой руки и зрительного восприятия обрабатываемого предмета, без появления центра, координирующего построение звуковых и визуальных образов. Еще одним условием функционирования языка, условием для возникновения речепроизводства является достаточно развитый спинной мозг, т. е. такой, который бы позволил достаточно тонко и точно управлять специальным режимом дыхания, обеспечивающим тонкие модуляции при подаче воздуха на голосовые связки. Все эти особенности появляются с достаточной определенностью только у кроманьонца, который появляется на исторической сцене лишь к концу периода мустье, т. е. к периоду 40 000 лет до р. Х. Все это делает сомнительными утверждения, которые относят время появления человеческой речи к двум миллионам лет до р. Х., к полутора миллионам лет до р. Х. и т. д. Явная же физическая асимметрия левого и правого полушарий головного мозга наблюдается только у неандертальца и кроманьонца. Ее появление относится специалистами лишь к периоду 30 000 лет до р. Х. При этом у неандертальца, по моим данным, положение надгортанника не располагает к звуковой речи. Из этого следует, что сама асимметрия конфигураций сильвиевой борозды в левом и правом полушарии у неандертальца свидетельствует, видимо, только о дифференциации в управлении правой и левой рукой и о появлении у него центра, координирующего движение правой руки и зрительного образа.
Таким образом, утверждения наиболее авторитетных авторов, цитируемых в замечательном по охвату семиотических работ труде В. Нёта (Nöth 2000), а именно Т. А. Себеока (1991, стр. 95), отнесшего время появления естественного языка к 2,5 - 3 млн. лет до р. Х., В. Коха, Дж. Уинда, М. Дональд (1991, стр. 122) относившей время возникновенния языка к 1,5 млн. лет до р. Х., в свете сказанного кажутся мне сомнительными.
1. 2. Animal rationale = animal symbolicum.
Так же как возникновение жизни на земле обязано возникновению первой на земле семиотической системы - генетического кода, появление в биосфере ее высшего, духовного начала - человека - обязано появлением у гоминида уникальной семиотической системы - человеческого языка.
Э. Кассирер, писал: «Великие мыслители, которые определяли человека как animal rationale (разумное животное), не были эмпириками, они не пытались дать эмпирическую картину человеческой природы. Таким определением они скорее выражали основной моральный императив. Разум - очень неадекватный термин для всеохватывающего обозначения форм человеческой культурной жизни во всем ее богатстве и разнообразии. Но все эти формы суть символические формы. Вместо того чтобы определять человека как animal rationale, мы должны, следовательно, определить его как animal symbolicum (символическое животное). Именно так мы сможем обозначить его специфическое отличие, а тем самым и понять новый путь, открытый человеку, путь цивилизации» (Кассирер 1998, с. 472).
Я согласен с Кассирером в том, что европейские ученые постоянно игнорируют огромную значимость того факта, что человек выделяется из среды прочих животных тем, что владеет самыми сложными и совершенными знаковыми системами. Однако с некоторой другой точки зрения можно сказать, что высокая степень разумности невозможна без овладения сложнейшими и наиболее прогрессивными знаковыми системами. Поэтому после установления связи между этими двумя свойствами можно уже будет считать, что вынесенные в заглавие характеристики - в некотором смысле тавтология. Во времена Кассирера в этом еще можно было сомневаться.
Ниже я изложу одну из точек зрения на связь между овладением сложными семиотическими системами и рассудком и затем - разумом в эволюции. Пьер Тейяр де Шарден в своей замечательной книге «Феномен человека» (Шарден 1987) неоднократно говорит о том, что история - это по существу развитие сознания, завуалированное морфологией. «С конца третичного периода, - пишет Шарден, - на протяжении более 500 миллионов лет в клеточном мире поднималась психическая температура. От ветви к ветви, от пласта к пласту, как мы видели, нервные системы, pari passu, все более усложнялись и концентрировались. В конечном счете, у приматов сформировалось столь замечательно гибкое и богатое орудие, что непосредственно следующая за ним ступень могла образоваться лишь при условии полной переплавки и консолидации в самой себе всей животной психики» (Шарден 1987, с. 139). Как же протекал этот процесс? Вначале нужно понять, что же было у живого существа до сознания, что руководило его поведением, что руководило адаптацией к меняющимся условиям жизни? Инстинкт. Но инстинкт это врожденная, геномиальная программа поведения. Следовательно, первым руководящим органом живого организма является геном5. Однако геном, как главный компонент управляющей системы неудобен тем, что он содержит в себе ограниченный набор программ адаптации к окружающей среде, который очень медленно изменяется, что влечет за собой слишком слабую, чтобы быть уверенным в том, что выживешь, общую адаптируемость организма к меняющимся условиям внешней среды6.
Ещё одно неудобство управления поведением со стороны "библиотеки" врождённых программ состоит в том, что в неё нельзя часто вносить изменения, поскольку от этого геномиальная книга портится. Уже поэтому геномиальная программа не может управлять изменчивым и вариабельным поведением особи, которая вынуждена лавировать между обстоятельствами, чтобы победить в конкурентном социальном общении, а может управлять лишь поддержанием тех или иных "несущих конструкций" в социальной системе вида путем принуждения особей к "правильному для вида" поведению, заставляя их удерживаться на стадии ритуализованных демонстраций вопреки желанию немедленно спариться или ударить противника (см., например, Фридман, 1992а, 1993а, б, в). Поэтому если человеческие слова выражают тот смысл, которые в них вложило ego, и в некотором смысле являются орудием мыслей индивида, при помощи которых он убеждает оппонента, то смысл сигналов животного - в указании ему и его оппоненту той позиции в видоспецифической социальной системе, которые они могут занять, если подчинят свои стремления и побуждения специфическому режиму исполнения соответствующей демонстрации, но отнюдь не являются орудием, позволяющим одному животному успешно конкурировать друг с другом в контексте агрессивного или сексуального общения (см., например, Фридман, 1998а, б; Фридман 1999; Friedmann, 1998).
Поэтому на некоторой стадии развития у живых организмов появляется новый инструмент управления - сложная нервная система, появляется условный рефлекс, а затем - зачатки сознания, которые позволяли организму все более и более точно учитывать и прогнозировать изменения окружающей среды и все более и более адекватно реагировать на них. Все эти изменения каждый раз становились возможными только благодаря усложнению рефлекторных комплексов и специализации знаковых систем, обслуживавших жизнедеятельность живых организмов.
Все живые существа, размножающиеся половым путем, вступают с партнерами по виду в коммуникативную связь. Коммуникативные системы не только поддерживают сообщества таких организмов в единстве, определяют границы вида7, но и структурируют эти сообщества и определяют его границы, как в пространственном отношении, так и в социальном. В частности они определяют не только размеры и конфигурацию территорий у отдельных членов группировки, но и структуру группировки в целом - набор рангов, которые могут иметь отдельные особи, допустимый успех их социального поведения, пропорциональный социальному статусу, напряжённость конкурентной борьбы за ранги и пр. В качестве примера действия подобной социальной системы можно привести скопления территориальных самцов на токах и территориальных пар в колониях у многих видов земноводных, птиц и млекопитающих. В такого рода скоплениях напряжённость конкурентного социального общения и степень социальной стратификации особей достигают максимума. В сообществах этого вида система угрожающих поз, распределенных с самим актом внутривидовой агрессии, как знаков, используемых для защиты своей территории позволяет особям определить демаркационные линии и поддерживать их (чем ближе к центру своей территории находится особь, тем более агрессивным по отношению к сопернику становится ее знаковое поведение), система сигналов при этом поддерживает ранговую иерархию особей в стаде.
У социальных животных геном отображает не только внутренние правила развития организма и его функционирования, но и правила социального взаимодействия, правила социальной организации, в частности, правила подачи сигнала и реагирования на коммуникативные акты собратьев по виду (и не только их), вначале - на сигналы воспитателя (отца, матери, родственников), затем - на сигналы вожака, на запахи своей стаи и т. д. В этом смысле можно сказать, что рефлексы, рефлекторные дуги, которые обычно рассматривают только в пределах организма, в случае коммуникации выходят за пределы тела одной особи и начинают действовать в пределах социума между двумя и более особями вступившими в рефлекторно-коммуникативный контакт: ответ на коммуникативный сигнал у низших организмов совершенно автоматичен, хотя и может быть выбран из библиотеки возможных ответов. Когда животному предъявляют релизер, оно может отказаться на него реагировать. Например, колюшка может не атаковать противника с красным брюшком, но если уж она реагирует, то реагирует единообразно; обязательность ответа на релизер есть обязательность единообразного поведенческого ответа на сигнал, как говорят биологи, высвобождающий (releasing) этот ответ, но не обязательность реагировать на сам сигнал.
В свете сказанного можно считать, что установление коммуникативного контакта на низших стадиях развития организмов является актом объединения двух и более организмов в единое рефлекторное целое, который позволяет рефлекторной дуге действовать в этом целом, как если бы она действовала внутри организма. Этологи уже получили доказательства того, что регулярные взаимодействия в коллективе особей (агрессивные или брачные) объединяют их в единую рефлекторную коммуникативную сеть (communicative network), заставляющую их координировать и синхронизировать не только моменты вступления в социальные контакты или ритмику перебора разных сигналов в процессе общения (хотя, кажется, что интересы каждой особи противоположны интересам всех прочих и могут быть реализованы лишь за их счёт), но и управляющую самим процессом выбора демонстраций. Один из механизмов, позволяющих видовой норме социальных отношений управлять поведением отдельных особей, выявлен в работах В. С. Фридмана (1993а - в, 1995а - г, 1998а, б), посвящённых анализу коммуникативных сетей в группировках больших пёстрых дятлов, охраняющих одиночные территории с августа по март следующего года.
Соответственно на ранних стадиях развития социальных структур тип коммуникативных систем, обеспечивающих жизнедеятельность социума, видимо, должен быть не просто адаптирован к геномиальному способу управления социумом, он должен быть и подобен тем коммуникативным системам, которые позволяют спинному и головному мозгу, а также подчиненным им ганглиозным узлам управлять организмом. Такой тип семиотических систем я буду называть геномиальным или автоматическим. Ядерный компонент такого рода систем передается по наследству и управляется инстинктами8. Приобретенная же в процессе жизни часть такого рода коммуникативных систем управляется условными рефлексами9.
Рассмотрим развитие примитивных коммуникативных систем на примере двух их типов, а именно релизеров и, так называемых, иерархических знаков, достаточно подробно рассмотренных В. С. Фридманом10. Впервые понятие релизера появляется в работах Хейнрота. Его формальное определение дано в классической работе Лоренца "Компаньон в мире птиц" (1935) и развито в работе "К понятию инстинкта" (1937). Термин РЕЛИЗЕР (AUSLÖSER) ориентирован на адресата. Для него это чистой воды стимул. И, если уж животное на него отреагировало, то его поведенческий ответ (несмотря на то, что он сложен и многоступенчат) никак не соотносится с внешними обстоятельствами (даже в том случае, когда он не достигает цели). Т. е. релизер для адресата коммуникативного акта служит специфическим (знаковым) стимулом, вызывающим инстинктивную реакцию (в отличие от стимулов, вызывающих рефлекторные реакции; они часто бывают неспецифичными и всегда непостоянны, тогда как инстинктивные реакции наблюдаются (в идеале) лишь в ответ на специфические стимулы (релизеры часто называют также SIGNI STIMULI - 'знаковые стимулы') и лишь тогда, когда животное находится в специфическом мотивационном состоянии. Регулярное социальное общение, без которого не могут существовать социумы на ранних этапах развития эволюции, необходимо (даже если оно происходит в агрессивном контексте) именно для того, чтобы поддерживать специфическую мотивацию, заставляющую животных не уклоняться от взаимодействий (и новых предъявлений релизеров), а искать их снова и снова, улучшая результаты прежней победы или компенсируя прошлые поражения (Tinbergen 1973). В понимании Н. Тинбергена релизеры - это знаки мотиваций. Лоренц в своей "Агрессии" фактически встаёт на эту точку зрения, хотя в его работе 1935 г. прослеживается более интересное понимание релизера как знака социальной роли данного компаньона в группировке. В. С. Фридман (1998а) понимает релизер, как знак потенциальной позиции животного в группировке, которую оно сможет занять, если общение, с использованием данного сигнала будет успешным. Доказательства того, что угрожающие и брачные демонстрации птиц могут передавать информацию о подобных возможностях, получены.
Итак, термин релизер был определен К. Лоренцем как стереотипный и демонстративный знак, который запускает стереотипную же последовательность актов инстинктивного поведения - комплекс фиксированных действий. В работах по коммуникации животных используются также термины триггер - от англ. trigger 'спусковой крючок' - и насос (см., например, Резникова 1997, с. 36, однако эти два последних термина можно использовать как гипонимы к термину релизер, в дальнейшем я так и буду поступать; при этом и под термином ТРИГГЕР, и под термином НАСОС я буду понимать такой тип сигнала, который требует автоматического ответа без возможности выбрать из библиотеки возможных ответов, каким именно сигналом нужно ответить и отвечать ли вообще; различие между триггером и насосом в принципе лишь количественное: реакция на триггер должна быть мгновенной, реакция на насос, грубо говоря, - когда кончится терпение; разница определяется тем, через какое время должен быть получен инстинктивный ответ на релизер, если мгновенно - то нужен триггер, если можно подождать, то насос гораздо надёжнее, т. к. обеспечивает более устойчивый результат. Я буду использовать термин РЕЛИЗЕР шире, чем это принято в биологии, поскольку не вижу принципиальной разницы в использовании его для обозначения знаков экстериорных систем и знаков интериорных систем, которыми, например, обмениваются клетки, хотя в отношении клетки термин ПОВЕДЕНИЕ требует некоторого уточнения. Есть еще и такая разница: в отличие от животного, реагирующего на релизер, клетка не может ни отказаться от реакции на гормональный или иной сигнал, ни выбирать какой-либо один сигнал из множества поступающих к ней. Такая свобода выбора существует при ответе на инстинктивные релизеры - в ответ на предъявленную противником угрожающую демонстрацию, например, дятел выбирает не ту, которая наиболее адекватна в смысле рефлекторного ответа на сигналы противника, а ту, которая с наибольшей вероятностью обеспечит победу для избранной ею тактики охраны территории (не только при данном, но и при другом виде ожидаемого сопротивления противника). Таким образом, общая тенденция в эволюционном развитии семиотических систем состоит в том, что либо увеличиваются возможности в выборе способов ответа на сигнал при фиксированном времени реакции, либо увеличивается возможность задержки ответа на сигнал. И в том и в другом случае развитие приводит к теоретически бесконечному числу возможных ответов или промежутку времени, за который нужно отреагировать на сигнал. И тот, и другой путь развития требует подключения к коммуникативному взаимодействию памяти и сознания, которое наполняет память моделями объектов и ситуаций, на которые нужно реагировать.
Однако различия между рассмотренными выше интериорными и экстериорными сигналами не выходят за пределы класса знаков, определенных Лоренцем указанным способом. Механизм действия релизерного знака, как инструмента запуска некоторой автоматической программы, и у клеток и у сложных организмов идентичен. Релизерные сигналы типичны для управления и клеткой, и тканью, и органом, и организмом. Примером интериорного триггера являются, например, гормоны. Так, стероидные гормоны диффундируют «сквозь плазматическую мембрану любых клеток, но только в клетках-мишенях они находят свой специфический рецептор, имеющий высокую степень сродства к гормону. Образуется комплекс гормон-рецептор, который далее подвергается «активации». <…> Гормон-рецепторный комплекс связывается со специфической областью ДНК и активирует либо инактивирует специфические гены. В результате избирательного воздействия на транскрипцию генов и синтез соответствующих мРНК происходит изменение содержания определенных белков, что сказывается на активности тех или иных процессов метаболизма» (Марри и др. 1993, т. 2, с. 159).
Типичным примером экстериорной релизерной коммуникации является демонстрация красного брюшка соседним самцом трёхиглой колюшки - самцу-владельцу территории вокруг "гнезда", который, восприняв сигнал-триггер, автоматически бросается в атаку. При этом он, как рецептор на определенный тип химического соединения, реагирует именно на красный цвет низа брюшка у рыбы, устремляющейся к построенному им "гнезду", а не на самого самца. Это было проверено на грубых моделях, практически не имеющих с обликом самца колюшки ничего общего.
Другое доказательство сходства между тем, как действует химический триггер, и тем, как действует животное, представляет пример с глухими индюшками, запротоколированный Вольфгангом и Маргарет Шлейдтами и воспроизведенный К. Лоренцом. Шлейдты сделали операцию на внутреннем ухе только что вылупившимся из яйца индюшкам, которые в результате этой операции оглохли. Когда они подросли, они посадили одну из них высиживать яйца. Когда из яиц вылупились и начали бегать вокруг наседки цыплята, она заклевала их насмерть. Лоренц и его коллеги объясняют это странное для матери поведение тем, что у индюшки сработал рефлекс, заставляющий ее отгонять от индюшат все, что движется и сравнимо с ее размерами, и не сработал рефлекс, тормозящий агрессию, поскольку он срабатывает только в ответ на писк новорожденных птенцов, а индейка была глуха. Визуально же она не может опознать в движущихся объектах собственных детей, поскольку «у нее нет ни малейшей врожденной информации о том, как должны выглядеть ее малыши. Она клюет все, что движется около ее гнезда и не настолько велико, чтобы реакция бегства пересилила агрессию. Только писк индюшонка, и ничто больше, посредством врожденного механизма включает материнское поведение и сдерживает агрессию» (Лоренц 1998, с. 141).
Релизерным может быть и акустический, и визуальный, и тактильный, и химический, и электрический тип сигналов. Важнейшей его характеристикой является то, что он - по терминологии Ч. С. Пирса является индексным и напрямую отражает мотивацию, т. е. поведенческую причину, автоматически запускающую сигнал. Примером релизерного сигнала у человека является непроизвольная улыбка младенца в ответ на склонившееся над ним грубое изображение человеческого лица, сразу переходящее в плач, как только мы изменим эти черты, придав им вид "лица" кошки (у других обезьян - еще и совы). У взрослых людей всё, что напоминает ответ на релизер по "ритуальности" и "инстинктивности", представляет собой двигательные стереотипы, обусловленные теми или иными социальными ритуалами.
У животных релизерные сигналы представляют собой чаще всего диалог, в котором инициатор-адресант подает инстинктивный сигнал партнеру по виду, на который партнер, распознавший в себе адресата, должен дать инстинктивный же и автоматический ответ.
Релизерные коммуникативные акты вызываются определенными типами мотивации11. Единая мотивация релизерных сигналов (например, защита собственной территории (концептуальный компонент знака)) объединяет их в замкнутую систему, в которой сигналы оказываются упорядоченными по силе побуждения (ценностный и одновременно энергетический компонент знака). Например, при защите территории, чем ближе к ее центру, тем более эффективную демонстрацию резидент (особь занявшая данную территорию и признанная сообществом особей ее владельцем) использует для ритуализованной угрозы захватчику, причём независимо от того, сосед это или новый вселенец. По мере того, как захватчик продвигается к центру территории и резидент пытается его остановить всё более эффективными демонстрациями, одновременно с каждой новой из этих демонстраций адекватно ей возрастает и то количество неритуализованной агрессии (клевков, выпадов, вытесняющих атак), которое резидент может предъявить захватчику одновременно с предъявлением ритуализованных демонстраций. Последние в этом случае в полной мере соответствуют понятию релизеров территориальной агрессии - чем выше их эффективность как средств территориальной коммуникации, тем большее количество неритуализованной агрессии может быть "высвобождено" и адресовано противнику у успешно удерживающей их птицы или животного (отметим, с одной стороны, обязательное сопряжение ритуализованной и неритуализованной территориальной угрозы, а с другой - их разделение во времени, позволяющее первой быть средством, а второй - орудием территориальной коммуникации).
«Таким образом, - пишет об этом К. Лоренц, - территория, которая, как кажется, принадлежит животному, - это лишь функция различий степени его агрессивности в разных местах, обусловленных локальными факторами, подавляющими эту агрессивность. С приближением к центру области обитания агрессивность возрастает в геометрической прогрессии» (Лоренц 1998, с. 87). Далее, однако, Лоренц показывает, что границы этой территории определяются взаимодействием или вернее противодействием конкурентов и противодействие это в слабой своей части воплощается в семиотическом диалоге, а в сильной - в потасовке. Последующие авторы (Hansen 1986, Popp 1987a, b; Фридман 1993а-в, 1998а, б) показали, что этот семиотический диалог представляет собой (равным образом как в контексте охраны территории или агрессивного доминирования, так и в контексте образования пары) последовательную эскалацию ритуализованной агрессии или ухаживания, когда птицы или животные всё быстрее меняют текущую демонстрацию на более эффективную и семантически богатую. При этом каждая из удерживаемых ритуализованных демонстраций одновременно оказывается и средством коммуникативной передачи более ценной информации, нежели предыдущий сигнал (в территориальной коммуникации это информация о возможности достичь более явной победы в стычке и сузить спектр действий, разрешённых подчинённым по отношению к данной птице или животному, см. Senar 1985, 1990), и орудием воздействия на поведение оппонента, позволяющим именно этой птице, а не её противнику, реализовать те "меньшие" возможности, передача информации о которых составляла семантику предыдущей демонстрации и реализация которых осуществляется в результате конкуренции обоих противников (Senar, 1990; Фридман, 1998; Payne 1998).
Тот диапазон силы специфического побуждения, который может быть отражён в семантике релизерных сигналов, может быть назван градиентом соответствующей мотивации (к охране территории, образованию пары, охране партнёра и пр., см. Фридман 1993б). Каждый из n сигналов-релизеров соответствующей мотивации выражает ограниченный отрезок этого градиента, не перекрывающийся с остальными. Число таких сигналов невелико (например, у изученного В. С. Фридманом большого пёстрого дятла их всего 8 в территориальной и 9 в брачной мотивации, см. Фридман 1993б, 1998а), и даже при наиболее дробном и интуитивном подходе к выделению демонстраций не превышает 15 - 20, хотя в указанной работе убедительно показано, что изменения уровня мотивации у взаимодействующих птиц или других животных происходят плавно, непрерывно, и гораздо медленнее, чем смена у них же демонстраций. Однако, с усилением системообразующей роли какого-либо из контекстов социального общения (охраны территории, агрессивного доминирования в стаях и пр.) для поддержания социальной организации вида на протяжении всего годового цикла, чем больше репертуар релизеров, "выражающих" данную мотивацию, тем богаче их семантика и "ýже" тот отрезок мотивационного градиента, знаком которых они являются (Фридман, 1993б; Иваницкий 1997). Эта закономерность позволяет видам, обладающим более специализированной охраной территории или более сложным процессом ухаживания, точнее нюансировать и детализировать собственные коммуникативные сообщения, благодаря чему у подобных видов исход социальных контактов оказывается гораздо более определенным и неоспоримым, формирующаяся при этом асимметрия социального поведения противников - более устойчивой и сильнее канализующей дальнейшее поведение обеих особей (Гольцман и др. 1994; Фридман, 1995а-в, 1998а, б).
Такое «неперекрывающееся расположение» релизерных сигналов на градиенте соответствующей мотивации, по мнению В. С. Фридмана (1993б, 1995) может быть уподоблено линейному расположению генов в хромосомах, в то время как цепочка демонстраций, последовательно предъявляемых резидентом в попытке остановить захватчика, прорывающего к центру территории - информационной РНК, прошедшей сплайсинг и готовой к «трансляции» - продуцированию социально ценных (повышающих устойчивость видовой нормы социальных отношений вопреки разнообразным возмущениям) действий, как по отношению к противнику, так и у самого противника12. Отмечу попутно, что в этих сравнениях системы релизерных сигналов с генетическим кодом есть скрытое указание на их подобие: по структуре своей релизерная коммуникация изоморфна действию генетического кода.
Релизерные сигналы в системе не могут действовать без дополняющего их другого вида сигналов. В. С. Фридман (1995) называет их эмоциональными. Они свободно комбинируются с релизерными, и при сопряжённом удержании тех и других в течение длительного времени они делают релизерный эффект неразличимым. Частота появления эмоциональных сигналов и степень их сопряжения во времени с соответствующими релизерами (особенно с наиболее эффективными сигналами, отражающими наивысший уровень агрессивной или брачной мотивации) возрастает тем более, чем дольше длится безуспешное взаимодействие противников, когда эскалация ими ритуализованной агрессии и переход к всё более эффективным демонстрациям, тем не менее, не создаёт устойчивой асимметрии в их сигнальном поведении и не позволяет выявить потенциального победителя. В. С. Фридман показал их наличие на примере общения при охране территории, но сходное дестабилизирующее воздействие на эффект релизеров при росте общего возбуждения птицы и соответственно - экспрессивности и экстравагантности вызванных ими действий и даже самих действий из состава демонстраций обнаружено у очень многих видов птиц (Панов 1978; Schleidt et al. 1984), и, видимо, эмоциональные сигналы представляют собой обязательную часть релизерной системы коммуникации.
Поскольку способность эмоциональных сигналов снижать эффект релизеров специфической мотивации прямо пропорциональна возрастанию опасности безрезультатного завершения длительного взаимодействия, то эмоциональные сигналы оказываются специфическими прерывателями релизерной коммуникации, прекращающими последнюю в тот момент, когда уровень агрессивного или сексуального возбуждения особей возрастёт до величин, угрожающих инвариантости коммуникативного восприятия соответствующих демонстраций и тем самым - их способности быть сигналами.
Эмоциональные сигналы, таким образом, необходимы при безуспешности взаимодействий, например, в случае, когда животные, равные по силе, зацикливаются в серии реплика - ответ на самой высокой ноте. Рост их представленности в потоке поведения у всех членов группировки, устойчивый рост доли прерванных ими социальных контактов оказывает дестабилизирующее воздействие на всю систему социальных отношений между этими особями; если оно продолжается достаточно долго, например, в результате постоянного притока всё новых вселенцев, претендующих на территории старых резидентов, то прежняя социальная организация разрушается, и социальная система вида в некоторых случаях переходит в новое состояние устойчивое к агенту прежней дестабилизации. Последнее характерно для случаев социального полиморфизма: полиморфизма социосексуальных систем у многих воробьиных птиц, в поселениях которых сосуществуют полигинные, моногамные и промискуитетные брачные отношения (Payne, 1983, 1985), для серии вариантов полиморфных территориальных отношений у большого пёстрого дятла (Фридман, 1993, 1995), поползня (Matthysen, 1986, 1989) и оседлых территориальных видов синиц-гаичек (Matthysen 1990; Ekman 1989).
Однако сложное поведение наиболее высокоорганизованных птиц (попугаи, врановые) и млекопитающих не может быть описано только с помощью представлений о релизерных механизмах коммуникации - её элементы, безусловно, сохраняются, но обслуживают уже более совершенный тип коммуникации, необходимый для поддержания более сложных социальных систем. В. С. Фридман (1993б, 1995) выдвинул гипотезу, что следующим этапом развития систем релизерной коммуникации являются системы, в которых коммуникация осуществляется с помощью сигналов, названных им иерархическими. Как и релизеры, они столь же эффективно выражают мотивацию животного, например, агрессивные демонстрации собаки или кошки столь же успешно выражают силу конфликта между стремлением нападать и бежать, что и демонстрации дятла. Однако если релизер передаёт информацию об объективно существующей в социуме позиции животного, которое оно может занять, если победит противника с помощью этого сигнала, то иерархический сигнал передаёт информацию о субъективном стремлении животного занять это место вопреки вероятному противодействию его оппонентов.
Это и позволяет иерархическим сигналам "перейти Рубикон" и стать средством отображения "личных" целей и планов, которые реализуются в поведении животного, и одновременно поставить устойчивость социальной системы в зависимость от успешности выполнения этих планов (вначале - лишь доминантами и подчинёнными, затем всеми остальными особями), тогда как при релизерной коммуникации социум управляет своими членами, а его устойчивость не зависит от исхода борьбы между стратегиями поведения разных особей. Очень красивые доказательства тому были приведены в работах А. Б. Керимова в соавторстве (1996) по большим синицам, где продемонстрировано, как в течение зимы из территориальной стаи избирательно удаляются те особи, чьё поведение не соответствует корреляциям, характерным для устойчивой популяционной системы этого вида (в частности, жёсткой корреляции между успехом особи в агрессивном доминировании и развитостью ряда морфоструктур, а также интенсивностью энергетического обмена), благодаря чему весной гнездовые территории занимают лишь особи, прошедшие этот отбор.
На основании сказанного выше можно сделать вывод, что у коммуникативной системы, состоящей из небольшого числа знаков, существуют лишь 2 возможных пути развития. Первый состоит в дальнейшей специализации системы релизеров, увеличении их числа, сопровождаемом сужением выражаемого ими «отрезка» мотивационного градиента. Обычно этот путь выбирается в случае успешной адаптации видов к такому образу жизни, при котором определённый тип социальных отношений - охрана территории у пёстрых дятлов, многолетнее поддержание брачных отношений в постоянной паре у гусей и пр. - оказывается всё более и более важным для поддержания всей социальной системы вида, однако сложность поведения каждой отдельной особи при этом в принципе остаётся без изменений, особенно в плане достаточно ограниченной возможности выбирать и направленно осуществлять долгосрочные стратегии социального поведения.
Второй путь состоит в преобразовании системы релизеров данной мотивации в систему иерархических сигналов, которые выражают ту же мотивацию, однако обозначают не ее уровень, наблюдаемый у данной особи в данный момент, а ту стратегию поведения, которую особь избирает для того, чтобы удовлетворить агрессивное или сексуальное побуждение данной силы в данном социуме, в условиях противостояния ему со стороны конкретных противников, у каждого из которых имеется определённый статус и определённые стратегии поведения. После такого преобразования знаковая система существенно усложняется и начинает сближаться в семиотическом отношении с человеческой речью сразу по нескольким независимым направлениям, каждое из которых усиливает все остальные.
Во-первых, сообщение, переданное иерархическим сигналом, может быть ложным - в случае, когда манифестируемая им стратегия поведения так и не осуществилась из-за противодействия противников, умиротворяющего поведения сторонников данной особи, или из-за того, что животное вообще не собиралось ее осуществлять. Исследование поведения обезьян (ср., например, Гудолл 1992) и даже псовых дает тому многочисленные примеры13, тогда как при использовании релизерной системы коммуникации все попытки социобиологов обнаружить "обман" при коммуникативном взаимодействии оканчивались неудачей. Особи не пытались предъявлять противнику демонстрации, не соответствующие их уровню агрессивной мотивации или возможностям победы. Даже искусственное повышение уровня их агрессивности (с помощью инъекций тестостерон-пропионата, см. Beletski et al., 1989, 1990) или перекрашивание, придающее им черты окраски доминантов, не вызывает ничего, кроме всплеска нападений на них со стороны всех членов группировки (включая даже подчинённых), которым они совершенно не способны противостоять.
Первый путь эволюции коммуникативных систем, по всей видимости, включает следующие процессы: «при сохранении старого набора мотиваций, но при необходимости их трансформации увеличивается число релизеров и сужаются обслуживаемые ими отрезки мотивационных градиентов. При исчезновении старых мотиваций и возникновении новых появляются новые релизерные комплексы <…> старые релизерные сигналы, переставшие быть эффективными в изменившейся системе социальных отношений, не исчезают, но начинают выражать не определённый уровень специфической мотивации, а возбуждение особи по поводу её неудовлетворения, т.е. становятся эмоциональными14, а простая перекройка пространства мотивационного градиента невозможна из-за однозначной связи между релизерной формой сигнала и его мотивационным содержанием» (Фридман 1995). И далее: «в эволюционной истории релизерные сигналы развиваются только по пути прогрессивной специализации, причем отработанные релизеры остаются в сигнальном фонде в виде эмоциональных сигналов. В результате общее число сигналов репертуара медленно, но неуклонно увеличивается, что ставит проблему отыскания преддемонстраций (действия из состава повседневного поведения, - незавершённые и начальные движения, движения намерения, смещённая и переадресованная активность, которая, в согласии с теорией ритуализации, служит эволюционным предшественником демонстраций-релизеров той или иной мотивации, см. Tinbergen, 1973; Хайнд, 1975» (Фридман 1995). Средства выражения, однако, оказываются вполне исчерпаемыми. Наступает кризис. Следовательно, релизерные системы коммуникации не могут в полной мере обеспечить ни развития, ни стабильного существования социума.
Релизерный тип знаковых систем, способный обслуживать тип социума лишь определенного уровня развития (понятие уровня здесь не связано с эволюцией, см. в этой связи интересные примеры, приведенные в книге Резникова 1997), имеет и другие ограничения. Так, релизеры имеют большую эффективность только в допустимых пределах оптимальной частоты взаимодействий. При выходе интенсивности взаимодействий в социуме за некоторые пределы релизеры резко снижают свою эффективность почти до нуля. Это ограничивает и тип социума, который этот вид коммуникативных систем может обслуживать. Он не может поддерживать социальные системы, которые не имеют определенного состава и стабильной плотности поселений. Как было показано Ж. И. Резниковой, к этому нужно добавить еще и пределы численности и плотности заселения сообщества. Так, в больших и плотно заселенных сообществах муравьев наблюдается более высоко организованная социальная структура, чем в относительно небольших, и одна и та же колония в процессе проведения опытов с увеличением численности муравьиного сообщества меняла социальную организацию и структуру. С этими изменениями связаны и изменения типов коммуникативных систем, которые поддерживают структурную организацию сообщества. Муравьи, взятые для опытов из малой муравьиной семьи, не проявляли тех коммуникативных способностей, которые были характерны для муравьев, взятых из большой и сложно организованной колонии (Резникова 1997, с. 56).
Кроме того, при релизерной системе коммуникации социальная система способна предвидеть и заблаговременно приспособиться только к таким изменениям среды, которые являются исторически типичными. При нетипичных изменениях выход может быть найден только эволюционным путем, который можно не успеть пройти.
Материалом для создания знаков и знаковых систем являются обычные жизненно целесообразные действия, которые В. С. Фридман называет преддемонстрацией (элементы повседневного поведения, эволюционные предшественники релизеров). Очередным шагом к превращению преддемонстрации в знак является демонстрация. Характерными признаками демонстрации являются, во-первых, утрированность отдельных элементов движения, переосмысление функции движения, преобразование его мотивации в знаковую, редукция и схематизация элементов движения, усреднение вариантов совершения движения, преобразование самого движения в его модель, наделенную другой мотивацией, другой функцией в программе поведения. Следующим шагом является закрепление самой демонстрации и столь же сигнификативно разработанного ответа на нее в геноме - образование безусловно-рефлекторного ритуала. Отмечу, что на низших стадиях развития экстериорной семиотической деятельности безусловно-рефлекторный ответ на коммуникативную ритуальную реплику является принципиально обязательным. Я связываю это с тем, что геном должен на этих ранних стадиях развития экстериорных коммуникативных систем действовать одинаково в интериорных и экстериорных коммуникативных системах. Для внутренних же коммуникативных процессов характерно, что базой проводки коммуникативного акта на высших уровнях коммуникативных систем является замкнутая рефлекторная дуга. Когда в коммуникативный акт вступают две особи, они с точки зрения рефлекторной деятельности на время коммуникации должны образовать как бы некоторое физиологическое целое. А коммуникативный акт должен представлять собой аналог замкнутой рефлекторной дуги, одна часть которой локализуется в организме адресанта, другая - в организме адресата. Эволюционным шагом в переходе от неконтролируемых рефлекторных знаковых действий к контролируемым является, на мой взгляд, процесс возникновения условно-рефлекторной привычки, обычая и ритуала, при котором цепь операций, образующих жесткий каркас рефлекторного действия, трансформируется, операция может, например, остановиться на каком-то неконечном этапе развития этого действия, и затем, эта первоначально как бы незамкнутая цепочка переориентируется на новую функцию, а именно, функцию нового релизерного сигнала, обычно также рефлекторного.
Следующий этап развития экстериорных коммуникативных систем может возникнуть только благодаря тому, что животный мир прошел этап релизерных коммуникативных систем. Более того, именно релизерные сигналы и преобразуются в сигналы нового типа, которые В. С. Фридман называет иерархическими по той мотивации, которую начинает отражать новый тип сигналов. Иерархические системы возникают из релизерных под воздействием того эволюционного тупика, в который их завела прогрессивная специализация. Поэтому и репертуар этих систем «будет складываться из релизерных сигналов той мотивации, на базе которой и для удержания которой в нестабильных условиях среды при сохранении общего консерватизма организации формируется иерархия» (Фридман 1995). В статье Фридман 1993 было показано, что при переходе от релизерной системы к иерархической основным иерархическим сигналом становится бывший наиболее эффективный релизер; если прежде разнообразие сигнального репертуара птицы или животного было составлено разными релизерами одной мотивации, то теперь его составляют разные варианты этого "основного" сигнала, изменчивость которых отражает колебания мотивации особи в отношении к соответствующему общению15. Прочие сигналы репертуара этой мотивации исчезают. Появление сигналов нового типа, видимо, совпадает с появлением нового типа социальной организации - иерархической. Появление же и коммуникативной, и социальной систем должно быть увязано с выходом на новую ступень биоинтеллектуального развития, что выражается в построении все более точных моделей окружающей среды и, соответственно во все более точном предвидении будущего. В этих условиях адаптации более широкого профиля сменяются на адаптации более узкого профиля, для чего требуются все более и более детализированные модели объектов окружающего мира. Только этим обстоятельством можно объяснить тот факт, что ранее запрещенные из-за нестабильности (точнее из-за неспособности поддержать стабильность социума релизерными коммуникативными системами!) типы социальной организации сменяют старые стабильные системы.
Иерархический тип социальной организации основан на том, что он вводит новый тип социальных отношений, более абстрактный, чем прежний. Если на предыдущем уровне организации социальной системы ее единство было основано на программах индивидуального контакта особей одного социума, то при иерархическом типе появляется отношение ранга, относящее всех индивидов к тому или иному ранговому классу (можно сказать, на индивида надевают погоны, показывающие, к какому рангу он принадлежит; вспомним в этой связи, как подписывался в официальных письмах Пушкин: «14 класса Пушкин»). Появляются таксоны, каждый член общества должен иметь теперь об индивиде новую информацию, основанную на личном узнавании16: к какому классу он относится. Далее следует процедура определения своего отношения к индивиду также через понятие ранга: ты выше по рангу или ниже, как строить свое поведение в отношении к данному индивиду становилось понятно из решения неравенства. Все это сказывается и на типах знаковых систем, которые начинают обслуживать новые социальные отношения. В новых типах знаковых систем между непосредственной мотивацией сигнала и самим сигналом встает новое звено - социальный ранг, который шифрует (представляет в свернутом виде) систему действий, которая будет осуществлена особью в условиях данной мотивации. Первоначально полученный в социальной системе ранг данной особи известен всему сообществу, соответственно, известной является и закрепленная за рангом последовательность действий в данной ситуации, остальное становится понятным из того, кто является источником сигнала. «Следовательно, - пишет далее В. С. Фридман, - смысл иерархического сигнала будет определяться не физиологией особи, но процессами, происходящими в социальной системе и его членением на ранги особей» (Фридман 1994). Я бы несколько изменил формулировку. Дело в том, что мотивация в случае иерархического сигнала не исчезает вообще и не исчезает из структуры знака. Ни один член социума не сможет обеспечить себе в нем успешной «карьеры», если он не понимает мотивацию, стоящую в данном случае за ранговой последовательностью действий. Изменение состоит в том, что к мотивации прибавляется еще один этаж означаемого знака и отношения между ближайшим компонентом смысла знака - рангом - и дальнейшим его компонентом становятся более сложными. Но такого типа знаки и рассчитаны на более изощренный тип интеллекта, чем тот, который опирается только на релизерные сигналы17. Все равно понимание знака предполагает вычисление мотивации, хотя в этом случае это не такой простой процесс, как ее «вычисление» в релизерном сигнале. Тем не менее, В. С. Фридман прав в том, что, по крайней мере, один из компонентов смысла знака вышел за пределы индивидуума в социум. Это знаменательный этап развития знаковых систем, с этого момента при возможности дальнейшего развития биоинтеллектуального моделирующего аппарата и коммуникативных систем (а это определяется сложностью нервной организации животного: например, у насекомых нервная организация достаточно примитивна, примитивен их «мозг» - ганглий, и их возможности ограничены, у млекопитающих она сложна, и у них мы и наблюдаем наибольший прогресс в этом направлении) создаются предпосылки для появления коллективных представлений. Отмечу здесь же, что с переходом к иерархическому типу коммуникативных систем изменились и все параметры коммуникативной системы, изменился и тип коммуникативного акта. Если в релизерных системах предложение коммуникативного контакта можно было рассматривать, как акт объединения двух простых организмов как бы в один сложный, то в иерархических коммуникативных системах установление коммуникативного контакта можно уже рассматривать, как только предложение отреагировать на сигнал. Особь должна сама каждый раз решать, реагировать на предложение или нет. В связи с этим над коммуникативной системой надстраивается система наказания за отказ от предложения, если предложение идет от более высокой по рангу особи. Релизерному сигналу такая надстройка не нужна.
Дальнейшее развитие, как социальной структуры, так и знаковых систем в мире животных связано с совершенствованием организмов в приспособлении к изменениям внешней среды. Приспособление к среде напрямую зависит от характера и точности ее моделей, а они в свою очередь зависят от характера и точности инструмента моделирования. Инструментом моделирования у живых организмов с момента появления на эволюционной лестнице нервной системы стали постепенно те ее «сгущения», которые располагались обычно в верхне-передних частях тела и служили первоначально только для реакции на стимул, потом - для управления жизнедеятельностью организма, потом - для моделирования объектов и ситуаций внешнего и внутреннего мира (предыдущие функции при появлении новых не отменяются). Чем больше и сложнее становились эти «сгущения», тем более точные модели был способен строить организм. Приведем примеры нервной организации живых существ от медузы и мухи до человека.
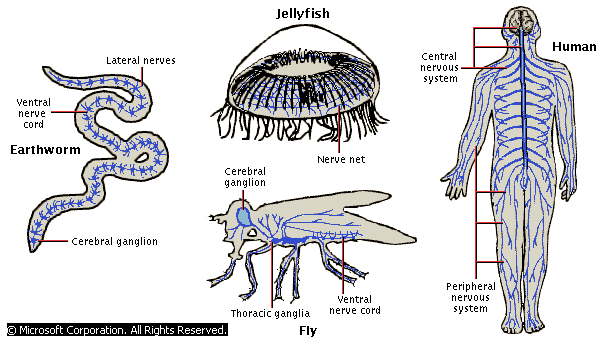
Нервная система появилась уже у низших животных как специализированная система клеток, действующая по схеме: стимул - реакция. В соответствии с этой схемой клетки нервной системы первоначально делились на сенсорные и моторные. В простейших организмах сенсорные клетки напрямую связаны с моторными, если, конечно не представляют собой две части одной и той же клетки. Скажем, у актинии, обладающей всего одной нейроноподобной клеткой, имеется нечто похожее на дендриты, воспринимающие стимул, и аксон, отдающий распоряжение мышечному волокну. «Что такая нервная система будет делать в ответ на стимул, столь же предсказуемо, как и то, что будет делать дверной звонок, если нажать на кнопку» (Наута, Фейртаг 1984, стр. 85). У медузы уже наблюдается нечто, подобное нервной сети. У нее два слоя клеток: сенсорные, находящиеся в прямом контакте со средой, и моторные, аксоны которых уходят к сократимым тканям, т. е. практически к мышечным волокнам. Следующий шаг в эволюции нервной системы был сделан уже теми же (но более сложно организованными) медузами. У них между этими двумя слоями нервных клеток появился новый, промежуточный слой, связанный не только с мотонейронами, но и друг с другом. Они задерживали ответ на стимул для того, чтобы соотнести его с информацией о других стимулах, упорядочить их и выбрать наиболее важный. Это уже элемент интеллекта и управления. Появление у червей ганглия навсегда решило вопрос о том, нужно ли управление. Уже у насекомых наряду с управлением появляется еще одна функция промежуточных структур клеток - моделирующая.
У мухи наблюдается уже разветвленная нервная система с единым нервным центром, который называется, как и у червя, церебральным (мозговым) ганглием (нервным узлом). Однако, он уже гораздо больше по объему и сложнее по устройству. Кроме церебрального у мухи есть еще грудной ганглий, напоминающий наш спинной мозг. Заметим, что у мухи несравнимо выросло и количество сенсуариев (под сенсуариями имеются в виду не только традиционные органы чувств, но и рассредоточенные по всему организму рецепторы типа болевых), пропорционально выросло и число ее интериорных и экстериорных знаковых систем. У высших позвоночных самая сложная нервная система, большой, развитый, сложно устроенный мозг, не отменивший и ганглии18. Мозг человека насчитывает порядка 1010 нервных клеток, из которых только порядка двух - трех миллионов мотонейронов. Сенсорные клетки вынесены за пределы мозга и располагаются в ганглиях. Все остальные клетки - элементы великой промежуточной сети, главным назначением которой является обработка информации, поступившей от сенсорных клеток и принятие решения о том, что делать с этой информацией (реагировать, отложить реакцию, запомнить, не реагировать). Мозг управляет организмом, и это касается как внутренней стороны его функционирования, так и внешней, которую можно подразделить на взаимодействие с природой и взаимодействие с себе подобными и не подобными. При этом управление организмом устроено не по принципу единовластия, а по принципу распределенности: мозг отдает общие команды низшим органам управления (грубо говоря, он знает, что нужно сделать), а те вводят в действие конкретные команды, которые должны выполнять конкретные телесные структуры (они знают, как нужно делать). Все сигналы, поступающие от рецепторов, многократно перекодируются и обобщаются. В мозг они поступают закодированными способом, удобным для мозга. То же происходит при передаче импульса от мозга к моторным клеткам. Сигнал проходит несколько стадий преобразований.
Основным назначением нервной системы является управление организмом. Само это управление можно разделить на управление частями организма и управление организмом как целым в его стремлении приспособиться к среде, в которой он живет. Всякое управление должно основываться на предварительном моделировании объектов, существенных для осуществления данной программы. Вот как об этом пишет К. Лоренц: «Второе понятие, к введению которого нас вынуждает изучение процессов приспособления, - это понятие знания. Уже в самом слове «приспосабливаться» неявно заключена предпосылка, что этот процесс устанавливает некоторое соответствие между тем, что приспосабливается и тем, к чему оно приспосабливается. То, что живая система узнает таким образом о внешней действительности, что в ней «отпечатывается» или «запечатлевается», - это информация о соответствующих данных внешнего мира» (Лоренц 1998, с. 263). Все это касается как внешних по отношению к организму объектов, так и внутренних. Для того, чтобы управлять жизнью клетки, необходимо иметь ее параметрическую модель, «знать» программы ее поведения, параметры ее состояний, опасные и безопасные отклонения от нормы в ее структуре и поведении. Точно так же для того, чтобы взаимодействовать с внешней средой, необходимо иметь модель объектов, релевантных для выполнения данной программы. Далее Лоренц пишет: «…приспособление - это возрастание взаимной информации (Transinformation) между организмом и окружающим миром. Такое возрастание происходит благодаря процессам, протекающим внутри организма, без заметного изменения при этом окружающего мира» (Лоренц 1998, с. 264).
Функцию моделирования релевантных для жизнедеятельности объектов выполняют в организме несколько приспособлений, между которыми, с одной стороны должно быть строгое распределение, с другой стороны, - перекрывание функций для надежности работы целого. Первым таким моделирующим аппаратом является геном, который на низших ступенях развития жизни делит эту функцию только с сенсорным аппаратом. Главные отличия между геномом и сенсорным аппаратом состоит в том, что 1) геном представляет собой хорошо организованное вместилище, предназначенное для накопления «знаний», причем оно изменяется каждый раз, когда к нему приращивается новый компонент «знания»; 2) «знания» хранятся в геноме в виде моделей объектов, программ, процессов, «сделанных» из одного и того же материала, а именно из комбинаций повторяющихся элементов - четырех оснований нуклеотидов аденина, тимина, гуанина, цитозина; 3) задача генома состоит в отборе правильных моделей и отбраковке неправильных; для достижения этих целей идет постоянное сопоставление модели и объекта, результатом которого является установление адекватности модели, кроме того, проверка правильности осуществляется и логикой построения самой модели; проверка идет долго и модель надолго же закрепляется в геноме. Сенсорный аппарат предназначен для представления постоянно меняющегося сиюсекундного состояния среды. В отличие от генома сенсорный аппарат в каждый данный момент должен быть сосредоточен на определенного рода задаче и в каждый следующий момент стирать все, что было запечатлено в предыдущий, и строить новую модель, подчас находящуюся в логическом противоречии с предыдущей. Таким образом, 1) сенсорный аппарат это устройство, принципиально не способное накапливать информацию; 2) модели сенсорного аппарата строятся на основе других принципов, чем в геноме, но между тем и другим способом моделирования должно быть установлено соответствие, позволяющее сверять результаты восприятия со «знаниями» генома; 3) в отличие от генома сенсорный аппарат должен строить не долговечные абстрактные модели, охватывающие классы случаев, а точные и конкретные, рассчитанные на миг существования модели.
С развитием нервной системы в организмах, ее обретших, происходит концентрация управления и долгосрочного моделирования в определенных местах, а именно в местах скопления нервных клеток, нервных узлов, ганглиев. Сформировавшийся у насекомых ганглий, преобразовавшийся у высших животных в мозг, стал центром, построенным в некотором отношении подобно геному: в нем хранилась и обрабатывалась информация, строились долгосрочные модели объектов, процессов, программ. По материалу и структуре они были более похожи на те модели, которые строил сенсорный аппарат, модели эти проверялись на адекватность быстрее и не столь тщательно и долго, мозг реагировал на быстрые изменения среды быстрее, чем геном, и это позволяло организму быстрее, с меньшими потерями и лучше приспосабливаться к среде. При этом, как отмечает В. С. Фридман, для успешного развития биоинтеллекта, для успеха в его конкуренции с геномом, «… для того, чтобы вызвать либо инстинктивный, либо рассудочный ответ на некий приходящий стимул (смешанный ответ существенно менее эффективен, как это показал еще К. Лоренц («Компаньон в мире птиц»)», Фридман 1995, стр. 14) центры, управляющие инстинктивным поведением и центры, управляющие «рассудочным» поведением должны быть разнесены по анатомически разным структурам. «Как известно, у млекопитающих такое разнесение имеется (рассудочные действия базируются в коре, а инстинктивные - в стриарных структурах), в то время как у птиц и те, и другие сосредоточены в стриатуме, хотя и в различных его отделах. Понять причину таких различий просто. В момент формирования птиц их предки - тероподы - находились в крупном размерном классе, и их более крупный мозг по законам аллометрии должен был управляться из геометрического центра, в то время как предки млекопитающих - терапсиды - находились в мелком размерном классе, и их более мелкий мозг мог управляться с периферии, из коры. Это вначале несущественное анатомическое различие позволяет птицам использовать рассудок только для оптимизации индивидуального поведения, в то время как млекопитающим - и для оптимизации социального поведения, в результате чего прогресс обучаемости в иерархических системах у птиц практически достиг своего предела, в то время как у млекопитающих этот потолок отсутствует, и данный прогресс уже дал нам человеческий разум» (Фридман 1995, стр. 14).
После появления у организмов органа, конкурирующего с геномом, дальнейшее развитие их характеризуется перераспределением функций между геномом и биоинтеллектом, ростом роли этого последнего вместе с массой мозга, увеличением времени, отводимого на воспитание потомства, усложнением социальной структуры живых существ, увеличением роли знаковых систем в управлении организмом, в функционировании социума как целого. Возникает союз четырех моделирующих механизмов: геномиального, сенсорного, биоинтеллектуального и знакового. Успехи в развитии и усложнении каждого их трех последних зависели от двух прочих.
Материал для моделирования поставляют в биоинтеллект сенсорные органы. Извлекают из моделирующего аппарата некоторые из этих моделей знаковые системы. Получается треугольник взаимно связанных и взаимно обусловливающих друг друга моделирующих систем, предназначенных для разных целей: сенсорная система, модели которой всегда связаны с моментом восприятия, поставляет информацию вовнутрь полностью внутренней биоинтеллектуальной моделирующей системе, модели которой принципиально не связаны ни с каким определенным моментом времени (это модели классов объектов, объектов вообще), а экстериорная знаковая система делает модели, построенные биоинтеллектом, доступными для социума. Модели знаковой системы животных, как правило, привязаны к моменту коммуникации, однако в иерархических системах в каждом знаке уже появляется абстрактный классификатор, интенсиональный компонент, приравнивающий друг другу все эквивалентные сигналы. Вся эта триада зависящих друг от друга моделирующих систем и эволюционирует системно: усложнение сенсорной системы позволяет усложнить биоинтеллектуальную систему, а эта последняя позволяет усложняться знаковой системе.
Внешним фактором для развития этой триады выступал социум. Зависимости здесь выстраивались таким образом, что, чем больше становилась доля рассудка в управлении социальной жизнью, тем устойчивее становился социум. Посредником в этом процессе должны выступить экстериорные знаковые системы, поскольку, как уже отмечалось, все социальные процессы, вся социальная структура держатся на знаковых системах, поддерживающих социальные функции19.
Усложнение знаковых систем поддерживалось, и со стороны означающего, и со стороны означаемого знака, и со стороны механизмов, поддерживающих связь между означающим и означаемым. По линии означающего - «Чем выше устойчивость социума, тем больше разнообразие характерных для него иерархических сигналов. Это требует прогресса рассудка животного в отношении предвидения свойств, характерных для этих сигналов (напомню, что иерархический сигнал - это лишь ярлык, а следующую за ним программу действия данного ранга животные должны понимать сами) программ социальных действий, что ведет к росту разнообразия сигналов, в свою очередь требующее дальнейшего прогресса рассудка. Таким образом, рассудочная деятельность и способность иерархической системы к предвидению прогрессивно развиваются, подстегивая друг друга» (Фридман 1995, стр. 15). Со стороны означаемого, как уже было отмечено, по достижении определенного уровня стабильности социума, дифференциация знаков пошла в направлении «трансформации смысла сигналов с отражения только программ социального поведения членов иерархического социума вначале в сторону отражения программ взаимодействия их с окружающей природой, а затем - и самих деталей окружающего мира» (Фридман 1995, стр. 16).
Дальнейший прогресс в развитии социума состоял в изменении нескольких важных параметров: 1) «с ростом роли рассудка в организации устойчивости и обучаемости социальной системы увеличивается степень преемственности и жесткость негенетической трансляции программ поведения в иерархической системе между поколениями. Одновременно с ростом жесткости трансляции по оси времени, в пространстве данная социальная структура приобретает способность сохранять свою устойчивость при все больших колебаниях состава индивидуумов в группах и все меньшей частоте их взаимодействий» (Фридман 1995, стр. 16); 2) нарастает степень свободы индивидуума от социума, а это означает, что геном все меньше координирует отношения между социумом и индивидом, все больше и больше обязанностей по координации этих отношений берут на себя социальные институты, правила социального поведения, особи, следящие за исполнением этих правил; такого рода перераспределение требует новых и новых знаковых образований, передающихся от поколения к поколению через воспитание; 3) возрастает роль индивидуального знания о природе, роль опыта и уменьшается роль физической силы социального доминанта20; 4) возрастает роль обучения, его сложность и время, которое на него требуется; 5) увеличивается набор социальных ролей, возрастает сложность организации социума и правил, по которым он живет.
Теперь приведу основной тезис В. С. Фридмана, касающийся развития знаковых систем: «… как только рассудочное поведение становится движущей силой обучаемости иерархического сообщества, а роль инстинктивного поведения становится пренебрежимо малой, то сразу же, по причине резкого усложнения социальной системы в ходе интенсивного роста обучаемости социума, требуется практически неисчерпаемый источник иерархических сигналов, не использовавшийся ранее инстинктивным поведением, поскольку в них следует вкладывать новый смысл, отсутствующий у уже имеющихся инстинктивных иерархических сигналов. В случае необнаружения такого источника прогрессивное развитие устойчивого социума средствами рассудка останавливается (поскольку его достижения необходимо выражать в сигналах - это является биологической основой того, что человек мыслит речевыми понятиями21) - на этой ступени остановились все млекопитающие, за исключением обезьян, у которых этот источник сигналов нашелся в виде богатейшей мимики (жирный шрифт - мой)…» (Фридман 1995, стр. 18). Полностью принимая этот вывод, я бы только хотел высказать некоторые сомнения по поводу утверждения, касающегося мимики обезьян. Для того, чтобы обрести систему знаков с неограниченным числом означающих, требуется комбинаторика, а каждый мимический сигнал у обезьян представляет собой отдельное сообщение и не комбинируется с другими сигналами в текст. Из отдельных мимических сигналов невозможно построить цепочки сигналов неограниченной длины. Для того, чтобы подобная система появилась, требовался совершенно иной тип организации, а именно, такой, чтобы наряду со структурно элементарными знаками в ней появились сложные знаки, чтобы сложность комбинированного знака была неограниченной, но чтобы сложность эта носила уровневый характер и чтобы на каждом из уровней системы количество комбинаций для построения сложного знака из простых теоретически было неограниченным. Если не считать сложно построенных ритуальных действий, такая знаковая система есть только у человека. Уровни сложности в ней суть следующие: фонемы образуют слоги и означающие морфов, элементарные компоненты смысла образуют означаемые морфов, из морфов строятся грамматические и фонетические слова, из грамматических слов строятся словосочетания, из фонетических - такты, из словосочетаний строятся предложения, из тактов - периоды, из периодов и предложений строятся сложные предложения, из сложных предложений - абзацы, главы, тома… Каждая языковая единица образует основу уровня комбинаторной сложности (фонетического, морфологического, лексического и т. д.). На каждом уровне строятся теоретически неограниченные цепочки исходных уровневых единиц. Следовательно, главными семиотическими проблемами эволюции знаковых систем при описании перехода от знаковых систем животных к естественному языку являются 1) физиологическая, которая разбивается на следующие а) какие инстинктивные ритуальные действия позволили гоминиду перейти к звуковой системе как базовой для образования языка?; б) какие физиологические процессы сопровождали этот переход i) в гортани; ii) в управлении системой дыхания; iii) в мозгу; в) какие изменения в семантике сигналов позволили сделать ее открытой; г) какие изменения в социальной структуре гоминидов позволили сделать ее открытой в смысле количества разнообразных ролей в общественной структуре; собственно семиотическая, которая разбивается на следующие подпроблемы: а) откуда в семиотической системе гоминида появились уровни; б) как произошел переход от комбинаторно простых к комбинаторно сложным знакам; в) каково происхождение морфологии, синтаксиса, сюжета. На некоторые из этих вопросов я постараюсь ответить ниже. Сейчас же на время перескочу ко времени, когда человек уже владел естественным языком, однако перед тем, как это сделать, подведу некоторые итоги этого параграфа.
Знаковые системы появились вместе с жизнью на Земле и, более того, инициировали саму жизнь (это положение еще будет обсуждаться в четвертой главе (второй главе второго тома).
Эволюция живых организмов - это «замаскированная морфологией» эволюция их эйдетического компонента: внутренней организации, функциональной структуры, сенсорного аппарата, знаковых систем, биоинтеллекта, социума.
Преобразование гоминида в человека связано с великой семиотической революцией - появлением у человека самой сложной и мощной семиотической системы - естественного языка.
Революция эта была подготовлена длительным эволюционным процессом, главными этапами которого явились а) переход от семиотических систем геномиального типа (релизерных) к иерархическим семиотическим системам, отделившим означающее знака от непосредственной поведенческой мотивации и поставившим между ними интенсиональный компонент; б) расширение репертуара означающих знаков; в) расширение круга именуемых объектов (областей референции и денотации или референтно-денотативных областей); г) переход от комбинаторно простых знаков к комбинаторно сложным; д) переход от одноуровневой структуры текста к многоуровневой; г) появление темо-рематического членения высказывания; д) появление сложно организованного сюжетом вербального текста.
Великая семиотическая революция была подготовлена координированным эволюционным развитием трех основных психофизиологических систем высших позвоночных - сенсорного аппарата, биоинтеллекта и знаковых систем, как интериорных, так и экстериорных. Внешним фактором, повлиявшим на развитие естественного языка, было координированное с развитием биоинтеллекта и знаковых систем прогрессивное развитие социума.
Семиотическая революция стала доминантным фактором для дальнейшего психического и социального развития человека.
Поскольку все семиотические системы у человека (экстериорные, интериорные, социальные, индивидуальные) составляют единое целое - индивидуальную семиосферу - все они (в том числе и интериорные моделирующие системы типа сенсорных), с одной стороны, влияют на формирование естественного языка, с другой стороны, начинают трансформироваться под его влиянием.