

Б, Допатоногве (Scaphopoda)
Дробление идет с образованием перед первыми делениями яйца полярной лопасти, связанной с*CD и с£>(рис. 78). Удаление полярной лопасти влечет за собой не только отсутствие у личинки производных 2d и Ы, но также теменной пластинки и прототроха. Изолированная бластомера CD дает нормальную, но маленькую личинку. Из яйца выходит настоящая трохофора (рис. 122,Л), у которой скоро, однако, появляется на спинной

Ряс. 122. Личинка и метаморфоз Dentalium (но Вильсону). А — трохофора; В — начало превращения; О — образование мантия; D — образование ноги и раковины.
стороне зачаток раковинной железы. Прототрох с еамого начала существования личинки состоит из трех колец крупных мерцательных клеток. Перед метаморфозом от раковинной железы вырастают две боковые складки, которые загибаютея к брюшной стороне и во время метаморфоза срастаются здесь, образуя трубчатую, несколько расширенную кпереди мантию, которая выделяет раковину такой же формы (рис. 122, С и D).
6. Головоногие (Cephalopoda)
По сравнению с яйцами остальных моллюсков (0,03—0,4 мм) яйца головоногих имеют громадные размеры (10—15 мм), так как в изобилии содержат желток, который составляет главную массу яйца. Протоплазма яйца в виде тонкого слоя занимает приблизительно У]2 поверхности яйца, так что объем ее приблизительно в 400 раз меньше объема желтка. Яйцо имеет почти эллипсоидальную форму, причем одиа из длинных сторон его несколько сплюснута (рие. 123). Протоплазма занимает область вокруг одного из полюсов яйца, и эта область отличается от желтка более светлой окраской. Положение развивающегося зародыша детерминировано и по отношению к сторонам будущего животного: анимальное полушарие яйца до дробления является дорзальным, вегетативное — вентральным, сплюснутая длинная сторона — задней стороной, а выпуклая, противоположная ей — передней стороной (рис. 123). Яйцо относится к телолецитальному типу, дробление идет дискоидально и не имеет никаких признаков спирального дробления, свойственного всем остальным моллюскам. Делится только протоплазма зародышевого диска, в желток же борозды дробления проникают на незначительную глубину
■136

е. по линии, идущей по меридиану от наиболее 3 о 8
только во время дробления, а затем все эмбриональные процессы идут в клетках диска, ютается пассивный. Норная борозда дробления делит зародышевый диск в направлении сагиттальной плоскости тела будущего зародыша, т. [уклон дл стороны к наиме-

н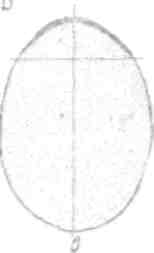 ее >й
(рис. L24). Вторая бо-
ее >й
(рис. L24). Вторая бо-
Перед»
Пр. Зад*
(а также проходит меридианально, перпендикулярно к плоскости первой' борозды, причем из получающв четырех бластомер две передние крупнее двух задних (рис. 124, В а 125, А). Также меридиа»
Рис, 123. Схематический разрез через яйцо Loligo pealii (по Ват аз). А — фр' I разрез; В
сагиттальный разрез; 0 — дорвальная сторона; в — вентральная; лев.—левая; пр.—правая; эадн.— н. ■ - передняя.
третьих борозды, причем они разде ляю! о !!.; двух передних бла стомер на две равные половинм, а. каждую из двух задних делят так, что, проходя параллельно первой бо розде п близко к ней, отделяют две [ькие бластомеры, лежащие в зад ней половине по бокам медиальной п.кк-кости (рис. 124, В и 125,4).
Толщина слоя, протоплазмы зародышевого диска настолько незначительна, что даже и после третьего и четвертого деления протяженность его протоплазмы остается наибольшей в направлении, параллельном поверхности яйца. В асе направлении ложатся и веретена четвертого и пятого делений, которые также разделяют диск в меридианальном направлении. При этом уже со второго деления группировка бластомер свидетельствует о билатеральности дробления (рис. 124 и 125), да и яйцо до дробления имеет форму, в которой может быть только одна плоскость симметрии.

Рис. 121. Начальные стадии дробления Loligo и npo- кже в них билатеральной л (по Нот а в).
g — Дораальная сторона.; Ь — центральна,!: лев.
девая; пр. —правая; пер. — передняя; яадн.— пяя.
Мо|шдт[аналг>щ,1о борозды у головоногих очень мало проникают в желток, и бла- сТОНбры первых четырех делений остаются соединенными с желтком своей перифери ческой частью; при пятом fee де лении, направленные к центру концы горых бластомер отделяются в виде свободных клеток от периферп- че, ких частей этих бластомер, сохра няющих соединение с желтком. То ясе происходит н при последующих деле- ниях дробления, тан: что центральное поле диска состоит на этих стадиях из постепенно увеличивающегося коли чества свободных мелких клеток, апе- риферия диска состоит из бластомер, дневных с желтком и пожучвв- шихназвание<1*а<#И0К0М0в(рис."126,.4.). Увеличение числа клеток централь ного поля происходит частью попе речным делением бластокоиов, частью раз iM самих свободных бла-
стомер, и вследствие этого диск постепенно расширяется по всем направлениям, а бластоконы смещаются нее дальше от полюса, сохраняя свое положение на периферия дни . Отделение новых бластомер от бластокоиов вскоре прекращается, и дальнейший рост диска обусловливается исключительно делением свободных клеток; с этого момента несколько расплывчатые очертания диска становятся гораздо более отчетливыми, а бластоконы обособляются от его краев и делением образуют группы клеток на желтке
. 137ч
у краев диска (рис. 126, В). Из этих клеточных групп на поверхности желтка образуется сплошпой эпителиальный слой, состоящий из сильно распластанных па я;ел-
„ тке
клеток. Его называют
тке
клеток. Его называют
jY J iy желточным эпителием,
или желточной энтодермой.
Ш"1
Рис. 125. Зародышевый
диск Sepia,
А —
на стадии 8-ми; В
— на
стадии
16-ти л
С —
па стадии 32-х бластомер; лев.
—
левая сторона
яйца; пр.—
правая; I—V
— направление первых-пяти делений
(I,
II,
III,
IV
и V
борозд деления) (по В и а л л ь т о л у).
--Г"
/*
2-4

■
■
..'Ч
Ш ■■■>
\?
 Рис.
126. Бластоднск Sepia
(по Виалльтопу). Слева—па стадии конца
дробления; справа—на стадии начала
образования зародышевых листков.
Рис.
126. Бластоднск Sepia
(по Виалльтопу). Слева—па стадии конца
дробления; справа—на стадии начала
образования зародышевых листков.
i — бластононы; S — бластомеры; 3 — желто»; 4 — утолщенпая часть бластодиска; б — тошна часть бластодиска; 6 — клетки, обособляющиеся от бластодиска.
После образования зародышевых листков мезодерма и эктодерма начинают быстро разрастаться непрерывным слоем по поверхности желточной энтодермы. В это время разрастаются также эктодерма и мезодерма в самом бластодиске, образуя в нем утолщения, складки и выросты, связанные с процессом формообразования.
138

А

Ряс. 127. Внешний вид зародыша Loligo (по Коршельту и Г с й д в р у) ца различных ранних стадиях развития.
1 — раковинный мешок; 2 — глазные выступы; з — винчение зачатка ; * — стомоЯеум; .5 — пояс вачатков щупалец; о — иооктодеум; 7 — зачаток жабры; s — отоцист; 9— зачаток мантии; т — задний зачаток воронки; 11— передний зачаток воронки.

Образование
внешней формы зародыша начинается
появлением в центре бдастодиска, т.
е. ванимальной или дорсальной части
зародыша, небольшого бугорка, верхушка
которого
образует впяче- ие—зачаток
раковин-нойжелезы(рис.127,А).
На несколько более Поздних стадиях края
этого винчения сближаются и срастаются,
так что раковинная железа образует
замкнутый мешочек и в виде мешка
разрастается по дорзальной стороне
но мере вырастания зародыша. Вслед
за появлс-н нем раковинной железы па
правой и левой частях бластодиска
об-равуютеявздутия вследствие
разрастания эктодермы.
Эти вздутия представляют собою зачатки
глаз и оптических ганглиев, причем сами
глаза появляются на дорзальной стороне
этих вздутий в виде впячений или глазных
ямок, замыкающихся затем в глазные
пузыри (рис. 127, В
и 0). На дорзальной же
стороне между глазными ямками возникает
в виде впячепия стомодеум, а у основания
бугорка раковинной железы закладывается
небольшой кольцевой валик — зачаток
мантии (рис. 128, 2).
ие—зачаток
раковин-нойжелезы(рис.127,А).
На несколько более Поздних стадиях края
этого винчения сближаются и срастаются,
так что раковинная железа образует
замкнутый мешочек и в виде мешка
разрастается по дорзальной стороне
но мере вырастания зародыша. Вслед
за появлс-н нем раковинной железы па
правой и левой частях бластодиска
об-равуютеявздутия вследствие
разрастания эктодермы.
Эти вздутия представляют собою зачатки
глаз и оптических ганглиев, причем сами
глаза появляются на дорзальной стороне
этих вздутий в виде впячений или глазных
ямок, замыкающихся затем в глазные
пузыри (рис. 127, В
и 0). На дорзальной же
стороне между глазными ямками возникает
в виде впячепия стомодеум, а у основания
бугорка раковинной железы закладывается
небольшой кольцевой валик — зачаток
мантии (рис. 128, 2).
|—7
Ъг-Н
Рис.
128. Послсдрвателыше стадии органогенеза
Loligo
(ориг.).
1
—
раковинный меток; 2 — маптия; 3
—
глав" ной
выступ; 4
—
жабра; 5
—
отоцист; в
—
попе щупалец;
7 — желточный мешок; 8—анальный
бугорок;
9—
задний зачаток воронки; 10
—
передний
зачаток воронки.
 ще
несколько позже раковинная железа
приподнимается над
поверхностью зародыша на
вершине бугорка, представляющего собою
зачаток туловища моллюска. Между ним и
глазными зачатками возникает еще
кольцевой валик — зачаток
воропки с парой отоцистов
у его основания, а на самом зачатке
туловища, па вентральной стороне,
появляется пара удлиненных
бугорков — зачатки жабер,
а между ними, по срединной
линии. — виячение проктодеума (рис.
127, С, 6). На
границе зародыша и желточного
мешка закладываются щупальца в виде
кольцевого ряда бугорков (рис. 128,6). На
следующих стадиях мантия нарастает
на туловищный бугорок и прикрывает
сначала раковинный мешочек, а затем
зачатки жабер и проктодеум;
на мантии появляются при
этом боковые плавпички. Зачатки
щупалец перемещаются к дорзальной
стороне головы и окружают стебелек,
которым зародыш соедипен
с желточным мешком (рис.
128, б). По мере закладки и разрастания
всех отих зачатков, последпие падавливают
па желток, который принимает в области
туловищного зачатка коническую суженную
форму, в то время как внезародышевый
желток остается грушевидным. На месте
соединения зародышевой и внезародышевой
половин желтка образуется резкий
перехват. Потребление желтка
идет наиболее интенсивно в задней части
зародыша,
ще
несколько позже раковинная железа
приподнимается над
поверхностью зародыша на
вершине бугорка, представляющего собою
зачаток туловища моллюска. Между ним и
глазными зачатками возникает еще
кольцевой валик — зачаток
воропки с парой отоцистов
у его основания, а на самом зачатке
туловища, па вентральной стороне,
появляется пара удлиненных
бугорков — зачатки жабер,
а между ними, по срединной
линии. — виячение проктодеума (рис.
127, С, 6). На
границе зародыша и желточного
мешка закладываются щупальца в виде
кольцевого ряда бугорков (рис. 128,6). На
следующих стадиях мантия нарастает
на туловищный бугорок и прикрывает
сначала раковинный мешочек, а затем
зачатки жабер и проктодеум;
на мантии появляются при
этом боковые плавпички. Зачатки
щупалец перемещаются к дорзальной
стороне головы и окружают стебелек,
которым зародыш соедипен
с желточным мешком (рис.
128, б). По мере закладки и разрастания
всех отих зачатков, последпие падавливают
па желток, который принимает в области
туловищного зачатка коническую суженную
форму, в то время как внезародышевый
желток остается грушевидным. На месте
соединения зародышевой и внезародышевой
половин желтка образуется резкий
перехват. Потребление желтка
идет наиболее интенсивно в задней части
зародыша,
139
СА
и на место желтка распространяется мезодсрмальпая ткань, из которой здесь формируется перикардии и сердце (рис. 129).
Мы видели, что эпителий, непосредственно одевающий желточный мешок, возникает из бластокопов, как желточная энтодерма; внутри самого вародыша желток одет кишечным эпителием, возникающим из краевого утолщения бластодиска. В одном небольшом участке втой кишечной энтодермы клетки собраны плотной группой, которая на более поздних стадиях отщепляется от эпителия желтка в виде плотного эпителиального мешочка. Этот мешочек представляет собою зачаток средней кишки, который затем растет в виде трубки по направлению к проктодсуму, другим же концом упирается в желток. После потребления той части желтка, которая находится на уровне анального отверстия) конец зачатка средней кишки растет несколько дальше и образует расширение, из которого формируется желудок (рис. 129, В, 17) и, кроме того, в виде выростов его возникают дольки печени. В то же время от стомодеума растет к этому расширению

прямая трубка — зачаток пищевода, который соединяется со средней кишкой (рис. 129, В, 15). Таким образом, не только энтодерма желточдого мешка, но и большая часть энтодермы, одевающей желток в зародыше, служит только для усвоения зародышем желтка и никакого участия в образовании кишечника по принимает; последний образуется из небольшого участка энтодермы зародыша и начинает pai ваться только после того, как обособится совсем от желтка.