

Глава 10. Корень
1. Происхождения корня и его функции
Корень, как и побег, возник вследствие морфологической дифференциации осевых органов (теломов) древнейших высших растений, обусловленной их жизнью на суше. Корням, как органу водообеспечения и минерального питания растения, дали начало подземные теломы —- ризомоиды, на поверхности которых развивались ризоиды (см. гл. 3, п. 1). В процессе эволюции корень возник, по-видимому, позднее побега. Осевое происхождение корня подтверждает и отсутствие существенных различий в анатомическом строении корня и стебля у некоторых видов высших споровых растений на ранних ет)вдиях их развития.
Из современных высших растений не имеют корней моховидные, поглощающие воду с помощью ризоидов, и псилотовидные, оба рода которых (псилот и тмезиптерис) имеют корневищеподобные органы в виде более или менее дихотомически разветвленных образований с многочисленными ризоидами. Морфологически они напоминают ризомоиды палеозойских риниевых.
Наряду с изначально («первично») бескорневыми растениями, среди высших растений встречаются и «вторично» бескорневые, утратившие корни в связи с водным образом жизни, как сальвиния из отдела папоротниковидных, пузырчатка, роголистник — из цветковых, или переходом к гетеротрофному питанию, как ладьян из семейства орхидных, встречающийся в болотистых местах северного полушария.
Корень, как и стебель, — радиально симметричный орган, характеризующийся апикальным ростом, осуществляемым деятельностью находящейся на его конце меристемы и участвующий в транспорте веществ.
Корень отличается от стебля отсутствием листьев, наличием чехлика, прикрывающего апикальную меристему, волосков, поглощающих из почвы воду с растворенными в ней минеральными веществами, и эндогенным ветвлением, то есть заложением зачатков боковых корней во внутренних тканях, удаленных от апикальной меристемы. Стеблю свойственно экзогенное заложение боковых почек, начало которым дают наружные части конуса нарастания побега.
Основные функции типичного корня:
Закрепление растения в субстрате.
Обеспечение водоснабжения и минерального питания растения, основу которого составляет поглощение из почвы воды с растворенными в ней веществами и проведение этих растворов в выше расположенные органы.
Участие в синтезе аминокислот, нуклеотидов, алкалоидов, гормонов роста и других физиологически активных веществ, образующихся при взаимодействии поступающих из почвы растворов с продуктами фотосинтеза, перемещающимися в корень из стебля и листьев.
Выделение ненужных метаболитов.
2. Морфологические особенности корня и корневой системы
Зачаток корня закладывается в процессе эмбриогенеза. У папоротниковидных он образуется сбоку от главной оси зародыша. У семенных растений зародышевый корень представляет собой продолжение оси будущего побега, так как он находится на противоположном конусу нарастания побега полюсе.
Граница между корнем и гипокотилем в зародыше неразличима. В развивающемся проростке ее можно определить по наличию на гипокотиле устьиц, которых нет на корне. Позднее между корнем и гипокотилем возникает перетяжка — корневая шейка.
При прорастании семени зародышевый корень углубляется в субстрат, закрепляя развивающееся растение. Этот корень, обладающий положительным геотропизмом, называют главным. У проростков он беловатый и часто почти прозрачный. Кончик корня конусовидный, довольно плотный и прочный, так как он должен противостоять давлению и трению о частицы почвы. На некотором расстоянии от кончика находятся корневые волоски. Они не только участвуют в поглощении воды, но и создают опору кончику корня, пробивающемуся между твердыми частицами почвы, способствуя, таким образом, его закреплению в субстрате.
По достижении определенной длины корень начинает ветвиться: зачатки боковых корней появляются выше зоны корневых волосков. Их развитие осуществляется акропетально: чем моложе боковой корень или его зачаток, тем ближе к кончику главного корня он находится, боковые корни растут более или менее горизонтально, так как в отличие от главного корня положительным геотропизмом они не обладают. Морфологически боковые корни сходны с главным корнем, но обычно тоньше и короче его. При их ветвлении образуются корни 2-го, 3-го и последующих порядков.
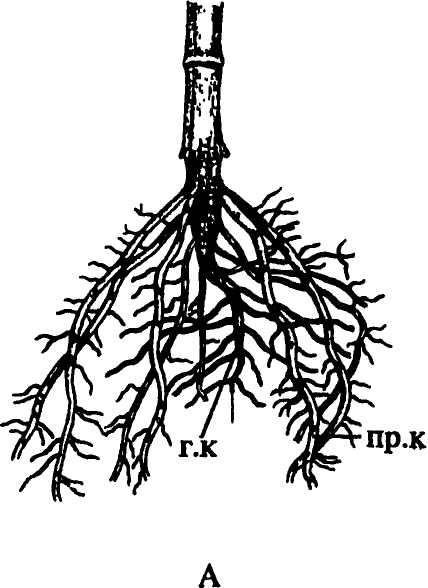
Зачатки боковых корней и развившиеся из них корни располагаются на главном корне продольными рядами — ортостихами, число которых варьирует у разных растений. У многих растений их 2, у гороха — 3, у грецкого ореха, дуба, ольхи — 6, у каштана — от 8 до 14. У однодольных число ортостих еще больше, но не всегда постоянное. Оргостихи могут отстоять одна от другой на одинаковое расстояние, как у тыквенных, молочайных, вьюнковых, у которых 4 ортостихи, но у гвоздичных, крестоцветных, зонтичных они попарно сближены. Число ортостих тесно связано с особенностями строения проводящей системы главного корня.
У взрослых растений боковые корни приурочены либо к верхней части главного корня, либо расположены в середине или ближе к его концу. Так как растения живут сообществами, это имеет важное значение, позволяя использовать воду и минеральные вещества из разных почвенных горизонтов.
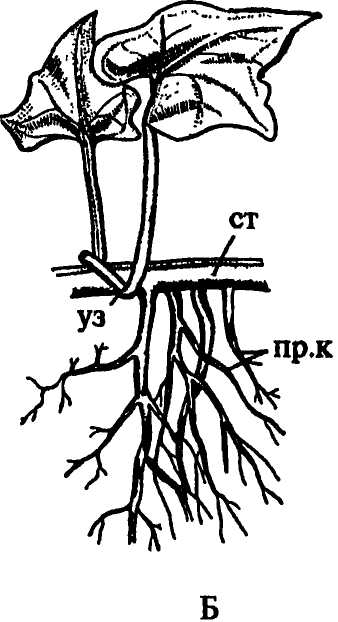
У некоторых растений очень рано появляются придаточные, или адвентивные (от лат. adventicius — пришлый, чуждый) корни, которые закладываются на гипокотиле, как у березы, недотроги, бальзамина, или на стебле.
Стеблевые придаточные корни в эволюции растительного мира возникли раньше главного. Они свойственны всем споровым архегониальным растениям.
Зачатки придаточных корней у них чаще всего закладываются близ апексов побегов. Их число и положение по отношению к листьям обычно постоянны для растений каждой систематической группы. У хвощей под листовым зачатком закладывается один корень, у папоротников — один или несколько, у плауновидных строгой приуроченности придаточных корней к листьям нет. Возможно, это связано с энационным происхождением листьев этих растений. Совокупность придаточных корней составляет гомо-ризную (от греч. homo — равный и rhyza — корень) корневую систему.
У семенных растений придаточные корни закладываются вдали от апексов побегов, число их не детерминировано и зависит от условий произрастания и наличия механических повреждений корней. Они приурочены к узлам, междоузлиям или пазушным почкам.
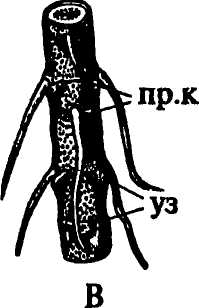
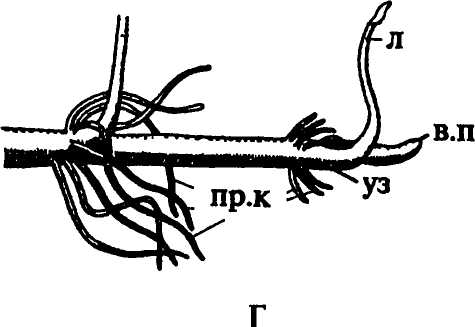
Наиболее часто встречаются узловые корни. Образование зачатков придаточных корней в узлах облегчается наличием в этих местах большого количества паренхимы, клетки которой не только долго сохраняют способность к делениям, но и содержат запасные вещества, расходуемые в процессе формирования придаточного корня. Большое значение имеет и наличие в узлах хорошо развитой проводящей системы, представленной не только пучками главного стебля, но также листовыми и веточными следами и их анастомозами, особенно обильными у однодольных растений. У мяты придаточные корни закладываются над и под узлами. У лютика, у представителей семейства зонтичных они развиваются под узлами (рис. 154 Г), у кукурузы и коланхоэ — над узлами (рис. 154А, В), у гвоздичных, пасленовых, многих розоцветных — в пазухах листьев. У плюща (рис. 154 Б), ириса, вороньего глаза, медуницы придаточные корни образуются на междоузлиях. У чистяка, лабазника, очитка едкого, кипрея их зачатки появляются на очень ранних стадиях развития побега, пребывающего еще в состоянии почки.
|
|
|
|
Рис. 154. Местоположение придаточных корней: А - надуаловые корни кукурузы; Б - междоузлиевые корни плюща; В - надузловые корни каланхоэ; Г - подузловые корни лютика. Обозначения: в.п - верхушечная почка, г.к - главный корень, л - лист, пр.к - придаточные корни, ст - стебель, уз - узел |
|||
|
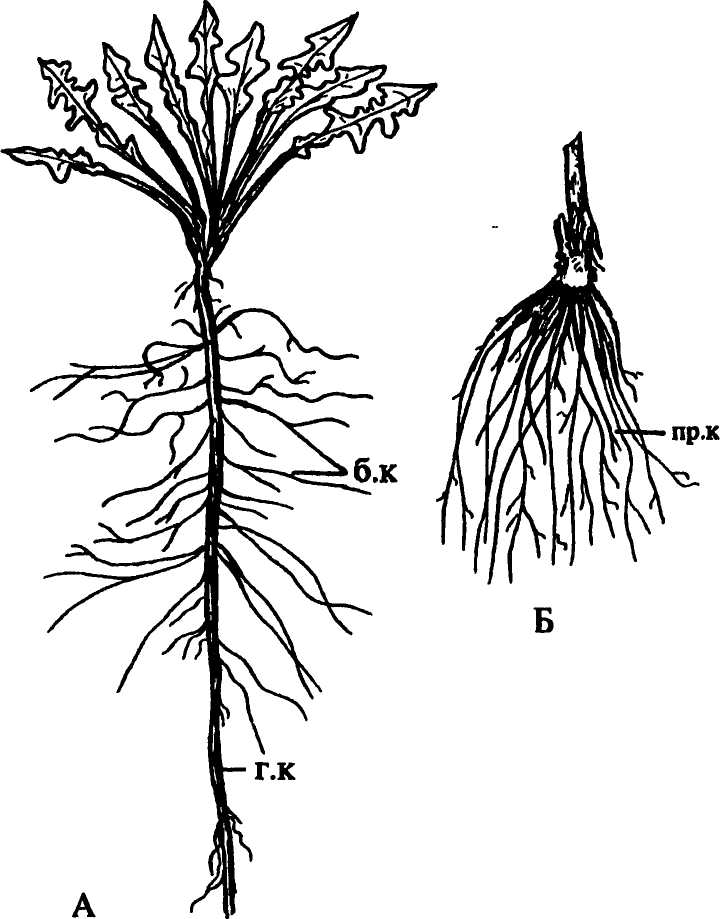
Аллоризная характеризуется сильным развитием главного корня, достигающего значительной длины и резко отличающегося от боковых корней. Такую корневую систему называют также стержневой (рис. 155 А). У некоторых растений главней корень, растущий вертикально вниз, достигает нескольких метров длины, у люцерны посевной, например, до 10-15 м. У большинства однодольных растений и некоторых двудольных (лютик, подорожник) главный корень быстро отмирает или степенью своего развития не отличается от многочисленных придаточных корней, составляющих пучок, или мочку. Такую корневую систему называют мочковатой (рис. 155 Б), или гоморизной, однако гоморизия у этих растений вторичного происхождения (в исходной гоморизии споровых архегониальных даже зачатка главного корня не бывает). У многих растений, преимущественно древесных — ели, сосны, дуба, березы, а также у некоторых травянистых многолетников, корневая система смешанного типа — с развитыми главным, боковыми и придаточными корнями. |
Рис. 155. Типы корневых систем: А — стержневая; Б — мочковатая. Обозначения: б.к — боковые корни, г.к — главный корень, пр.к — придаточные корни |
Интенсивность развития корневой системы тесно связана не только с видовыми особенностями растений, но и с внешними условиями. Так, у мятлика однолетнего, растущего на тяжелых глинистых почвах, корни достигают длины 10 см, а у растений, встречающихся на прибрежных песках, — 25-30 см.
В пределах корневой системы корни функционально дифференцированы. Особенно четко это проявляется у древесных растений. Длинные долговечные корни, служащие для расширения площади питания растения и его закрепления в почве, называют ростовыми, или скелетными. Образующиеся на них недолговечные короткие корни, осуществляющие поглощение воды с растворенными в ней минеральными веществами, называют питающими, или сосущими. Они регулярно развиваются весной на скелетных корнях и к осени обычно отмирают.
|
Сосущие корни способны вступать в симбиоз с грибами, называемый микоризой (от греч. mykes - гриб). Термин «симбиоз» предложен А. де Бари в XIX в. для определения одного из видов сожительства организмов, при котором они получают взаимную выгоду в осуществлении питания и водообеспечения. В настоящее время этот тип взаимоотношений называют мутуализмом. Гифы гриба либо снаружи оплетают корень (такую микоризу называют эктотрофной, или эктомикоризой), либо они проникают вглубь корня и живут в его клетках, где сильно ветвятся, образуя клубок (такую микоризу называют эндотрофной или эндомикоризой). В некоторых случаях сочетаются оба типа микоризы. |
Рис. 156. Обычный (А) и микоризный (Б) корни ели |
Микоризные корни не имеют корневых волосков. Их апикальный рост рано прекращается, а необычное ветвление приводит к образованию своеобразных утолщений (рис. 156).
Питание растений, осуществляемое с помощью гриба, называют микотрофным. Гриб разлагает некоторые находящиеся в почве минеральные соединения, способствуя, таким образом, их усвоению растениями. Из этих веществ особенно важное значение имеют соединения фосфора и азота. Увеличивая концентрацию клеточного сока, минеральные вещества повышают сосущую силу клеток, усиливая поступление в корень воды. Благодаря ферментативной деятельности, гриб гидролизует находящиеся в клетках корня углеводы. Часть их идет на питание самого гриба, а неиспользованные углеводы поступают в клеточные вакуоли, способствуя, как и минеральные вещества, увеличению сосущей силы клеток. Гриб способен также поглощать воду даже из относительно сухой почвы. Таким образом, микориза осуществляет не только минеральное питание, но и водоснабжение растений. Микоризы защищают корни от заражения их патогенными организмами. Используя углеводы, поступающие в корень по нисходящему току, микоризные грибы не дают им возможности нормально развиваться в клетках растения-хозяина, а вырабатывая вещества типа антибиотиков, они способствуют их уничтожению.
Не случайно поэтому микориза широко распространена в природе. Она отмечена более чем у 70 % голосеменных и покрытосеменных, а также у 60 % высших споровых растений. Некоторые цветковые растения, например орхидные, вообще не могут развиваться без микоризы. Не образуют ее представители семейств крестоцветных, маковых, крапивных, осоковых и других. Она не обнаружена у мхов, а также у насекомоядных и паразитических цветковых растений.
|
У растений, главным образом у бобовых, встречается и другой тип симбиоза - с азотофиксирующими бактериями из рода Rhyzobium. Растение реагирует на их внедрение активными делениями клеток молодого корня, приводящим к образованию опухоли, имеющей вид клубенька (рис. 157). Живущие в нем бактерии ассимилируют молекулярный азот, переводя его в соединения, легко усваиваемые растениями, которые, в свою очередь, обеспечивают бактерии органическими питательными веществами. По сравнению со свободно живущими азотофиксирующими бактериями, затрачивающими 50-100 и более граммов углеводов на фиксацию 1 грамма азота, бактерии-симбионты очень экономны и используют всего 3-4 грамма углеводов на 1 грамм азота. Фиксированный клубеньковыми бактериями азот остается в растениях, поэтому они успешно растут на почвах, бедных доступными для них формами азота. Имеется много рас клубеньковых бактерий, и не все они характеризуются высокой азотофиксирующей способностью. Образование многочисленных мелких клубеньков на боковых корнях вызывают обычно малоэффективные расы этих бактерий. Наиболее энергичный процесс усвоения азота происходит внутри крупных клубеньков, развивающихся на главном корне. Не все бобовые способны к симбиотическим связям с клубеньковыми бактериями. Из 13 000 видов этого семейства клубеньки на корнях обнаружены примерно у 10% видов. |
Рис. 157. Клубеньки на корнях бобового растения: А - расположение клубеньков; Б - поперечный срез клубенька. Обозначения: б.к - боковые корни, б.тк - бактероидная ткань, г.к - главный корень, кб - клубеньки, п.к - первичная кора корня, п.п - проводящий пучок, б.тк - бактероидная «ткань» |
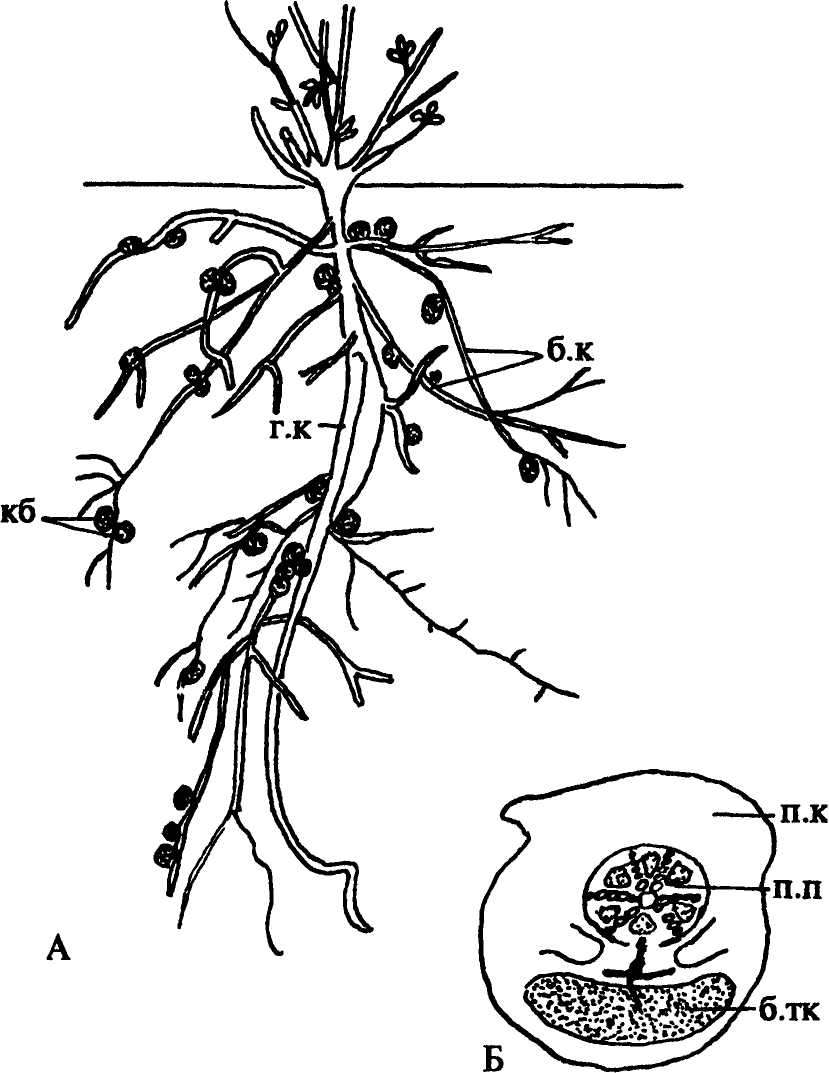
Образование корневых клубеньков может быть вызвано и другими бактериями, способными фиксировать молекулярный азот. Такие клубеньки характерны для корней ольхи, кофейного дерева, вейника лесного и других растений.