

7 Фізіологія сенсорних систем
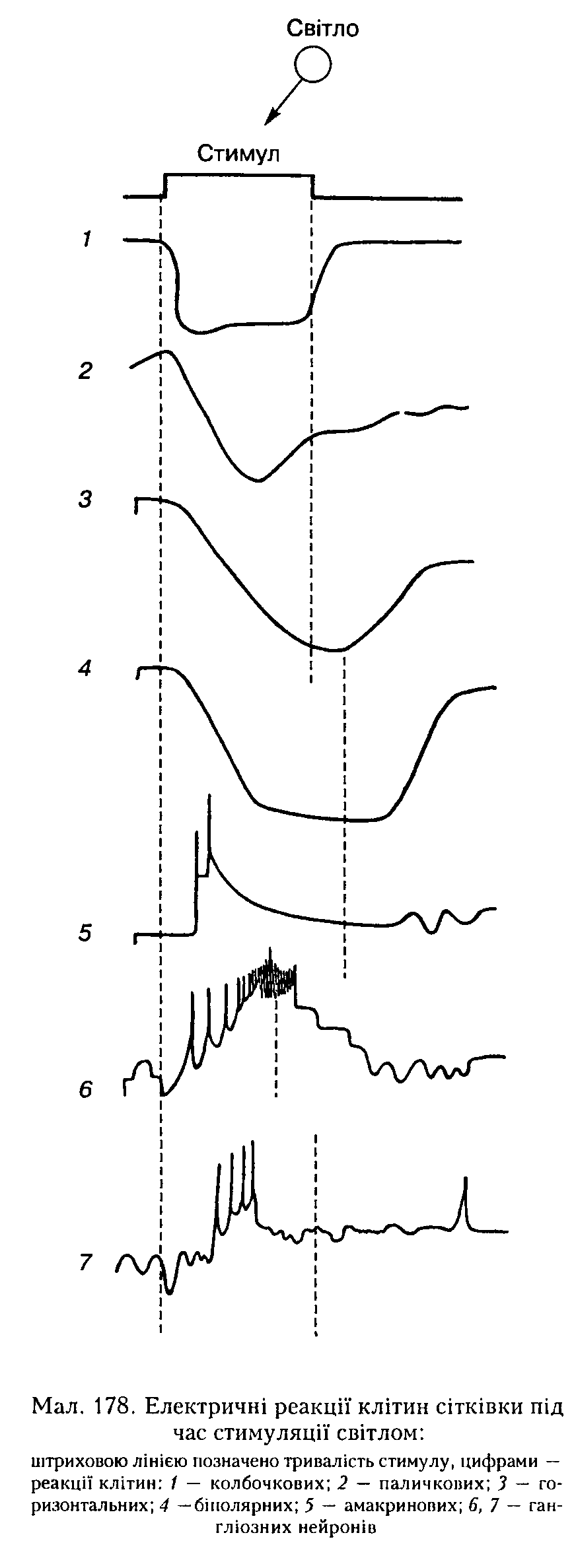
(нейрони С) реагують гінер- або деполяризацією залежно від довжини хвилі подразнювального світла.
Рецептивні поля гінгліозних нейронів. Функцію гангліозиих нейронів вузлового (гапгліозного) шару сітківки вивчають, реєструючи електричну активність окремих волокон зорового нерва. На кожному такому нейроні коивергує величезна кількість інших нейронів сітківки. Шляхом посилання дуже вузького пучка світла па різні точки сітківки можна знайти ту її ділянку, подразнення якої світлом спричинюватиме збудження або гальмування певного гіигліозиого нейрона. Ця ділянка і є його рецептивним полем. По-перше, гаигліозні нейрони реагують па освітлення сітківки короткочасно: лише на вмикання світла (нейрони-оп), лише па вимикання (нейрони-оЦ) і па вмикання й вимикання світла (нейрони-оп-о({). По-друге, нейрони рецептивного поля, розміщені в його центрі й на периферії, реагують на освітлення протилежним чином: якщо у відповідь па освітлення центра рецептивного поля иейрон-оя збуджується, то під час дії світла па периферичні частини цього поля цей нейрон гальмується. Відповідно він гальмується па вимикання світла в центрі рецептивного поля і збуджується па його периферії. Описане явище дістало назву бічного гальмування, воно зумовлене реципрокними взаємозв'язками між центральними і периферичними нейронами рецептивного поля.
Електроретинограма. У 1859 р. Е. Дюбуа-Реймон встановив, що між передньою і задньою частинами ока існує різниця потенціалів, яка змінюється під час освітлення. Запис цієї сумарної відповіді сітківки па освітлення називається електроретинограмою (ЕРГ) і складається з кількох коливань (хвиль) потенціалу (мал. 179), які відображають різні процеси: а-хвиля — початкове негативне коливання, зумовлене сумацією РП* фоторецепторів і горизонтальних нейронів; велика позитивна б-хвиля, зумовлена активізацією гліальних клітин сітківки; тривала позитивна в-хви-ля відображає зміни МП пігментоцитів —
Фізіологія зору 9

клітин пігментного шару сітківки на вмикання світла і г-хвиля — на вимикання світла, або так званий о/'/'-ефект. Та обставина, що ЕРГ відображає активність більшості елементів сітківки, дала змогу використовувати цей показник в офтальмології для діагностування деяких очних хвороб.
Світлова і темнова адаптація. Якщо загальне освітлення змінюється, то зорова система пристосовується до нових умов, змінюючи свою чутливість. Коли людина виходить з яскраво освітленої кімнати на темну вулицю, то в перший момент вона не може розрізняти навколишні предмети. Проте згодом їх контури стають помітними. Під час цієї темпової адаптації абсолютна чутливість зорової системи повільно зростає, і максимальної чутливості вона досягає лише через дві години перебування у повній темряві. Абсолютна чутливість у цьому разі становить (у розрахунку на один рецептор) 1—4 фотони світла за 1 хв.
Протилежний процес називають світловою адаптацією. Вона відбувається значно швидше, протягом кількох секунд (хоча на короткий час людина може бути повністю засліплена).
Вирішальну роль у процесах адаптації відіграють нейроииі механізми, які перемикають колбочковий зір на паличковий і навпаки. Крім того, важливим механізмом темпової і світлової адаптацій є зміна діаметра зіниці, що відбувається під час зміни рівня освітленості ока.
Й. В. Гете, який був не лише видатним поетом, а й природознавцем, описав таке явище: "Я повернувся до готелю повечеряти, і до моєї кімнати увійшла й стала на певній відстані від мене пишна покоївка з слінучо-білим обличчям, чорним волоссям і в червоній сукні. Я уважно придивився до неї. Після того як вона вийшла, я побачив на білій стіні проти себе чорне обличчя, оточене світлим пасмом, а одяг цієї цілком нової фігури видавався чудового синьо-зеленого кольору".
Це явище пояснюється локальною адаптацією і виникненням послідовних образів. Локальна адаптація виникає тоді, коли за сталої середньої освітленості обмежена ділянка сітківки має освітленість, яка відрізняється від середньої. Так, якщо протягом ЗО с фіксувати зір на якомусь зображенні, потім перевести погляд па рівний темний чи світлий фон, то протягом кількох секунд можна бачити негативний послідовний образ. У цьому послідовному образі те, що було па зображенні темним, видаватиметься світлим і навпаки. Чому так відбувається?
Ті ділянки сітківки, на які припадають темні місця зображення, за час фіксації зору стають чутливішими, ніж сусідні ділянки, на які припадали світлі місця зображення. Такі послідовні образи зберігаються досить тривалий час, якщо застосу
вати сильне освітлення сітківки або якщо "засвічування" (не дуже сильне) триває довго. У разі локальної адаптації до колірних стимулів утворюються послідовні образи, забарвлені у додаткові кольори.
У 1825 р. Ян Пуркіньє помітив, що вдень видаються яскравішими червоні кольори, а в сутінках — сипі. Це зрушення Пуркіньє пояснюється переходом від фото-нічного до скотопічпого зору під час темпової адаптації.
12.2.4. КОЛІРНИЙ ЗІР
Коли світло надає на певний об'єкт, то найчастіше відбувається один з трьох процесів: 1) світло поглинається і його енергія перетворюється па теплоту; 2) світло проходить крізь об'єкт; 3) світло відбивається від поверхні об'єкта. Часто можуть відбуватися два або три процеси одночасно. Для багатьох об'єктів відносна кількість світла, що поглинається і відбивається, залежить від довжини хвилі. Так, зелений листок рослини поглинає довгі й короткі хвилі сонячного світла і відбиває світло проміжної ділянки спектра, що сприймається нами як зелений колір.
Проте який саме колір ми бачимо, залежить не тільки від довжини хвилі, а й від розподілу енергії (інтенсивності) між різними частинами спектра і від неоднакової стимуляції фоторецепторів колірного зору — колбочкових фоторецепторів.
Теорії колірного зору. З погляду трикомпонентної теорії кольоросприйнят-тя Юнга —Гельмгольца, у сітківці є три

види колбочкових рецепторів, які функціонують як незалежні оптичні приймачі. Дійсно, ири вимірюванні мікроспект-рофотометром поглинання променів різної довжини одним колбочковим рецептором виявилося, що є колбочкові фоторецептори, які максимально поглинають червопо-оранжеві (560 нм), зелені (530 пм) і сипі (430 нм) промені(мал. 180).
Отже, в сітківці виявлено три групи колбочкових фоторецепторів, кожний з яких реагує па промені одного з трьох основних кольорів видимого спектра.
Зоровий пігмент колбочкових фоторецепторів йодопсин об'єднує три пігменти — еритролаб, хлоролаб і ціанолаб. Кожен з них має максимальну спектральну чутливість, що відповідає трьом основним кольорам спектра. Справедливість трикомпонентної теорії кольоросприйнят-тя підтверджується також законами змішування кольорів і багатьма психофізичними даними.
Електрофізіологічні дослідження показали, що електрична активність деяких гангліозиих нейронів шару сітківки виникає під впливом проміння будь-якої довжини хвилі у видимій частині спектра. Такі елементи сітківки називають домінаторами (Р. Граніт). Одні доміпатори отримують сигнали від паличкових фоторецепторів і називаються скотопічними, інші — від колбочкових і називаються фотопічними.
В інших гангліозиих нейронах, що називаються модуляторами, ПД виникають лише при освітленні їх промінням певної довжини хвилі. Виявлено 7 видів таких модуляторів, які оптимально реагують па світло різної довжини хвилі (від 400 до 600 нм). Три компоненти кольоросприй-пяття виникають унаслідок усереднення кривих спектральної чутливості модуляторів, які можна згрупувати відповідно до трьох основних частий спектра: сипьо-фіо-летової, зеленої, жовтогарячої.
У XIX ст. Герінг запропонував теорію опонентних кольорів, відповідно до якої існує 4 основних кольори — червоний, жовтий, зелений і синій. Ці кольори з'єднані попарно за допомогою двох антагоністичних механізмів: червоно-зелено
Фізіологія
зору
го та жовто-синього і називаються опо-нентними, оскільки в певний момент збуджується тільки один колір з кожної пари. Отже, теорія Геріпга постулює наявність антагоністичних иейроиних механізмів: якщо нейрон збуджується під виливом зеленого стимулу, то червоний стимул викликає його гальмування.
Сучасні мікроелектродиі дослідження показали, що у деяких хребетних, що мають колірний зір, існують "червопо-зелепі" нейрони, в яких виникає гіиерполяризація, коли його рецептивне поле освітлюється короткохвильовим світлом у діапазоні 400 — 600 нм, а при освітленні світлом довжиною хвилі понад 600 нм виникає деполяризація мембрани. Припускають, що такі нейрони найефективніше кодують інформацію про колір. Справедливість теорії оноиеитних кольорів підтверджується й наявністю кольорових послідовних образів.
Отже, під час дії на фоторецептори світла певної довжини хвилі виникає відповідна комбінація збуджень колбочкових фоторецепторів , яка визначає комбінацію збуджень у системі оноиеитних нейронів і створює максимум збудження па одному з селективних детекторів кольору. Як наслідок, виникає селективне і суб'єктивне сприйняття кольору.
Колірний просторовий зір розв'язує два основних завдання: опис кольору (барви) предмета чи його частин і впізнання предмета за допомогою кольору. На світлове випромінювання певної довжини хвилі колбочкові фоторецептори трьох різних тинів відповідають сигналами, які відрізняються за своєю амплітудою. Якісно ж усі сигнали однакові і колірних відчуттів у собі не несуть.
У проекційній зоровій зоні кори півкуль великого мозку існують рецептивні поля, які реагують на забарвлені певним чином розміщені смужки, а також звичайні концентричні колірні поля. Психофізіологічні дослідження свідчать про те, що в зоровій зоні кори мозку людини існують чер-вопо-зелені і синьо-жовті канали, які чітко відрізняються від ахроматичних. Надзвичайно важливою особливістю зорової системи є можливість пізнавати забарвлення предметів незалежно від умов освітлення. Це явище називають константністю кольоросприйняття. Воно зумовлене наявністю у зоровій зоні кори нейронів, які реагують у вузькій смузі частот незалежно від спектра, відбитого від поверхні об'єкта.
Крім спектральних кольорів існують і неспектральні кольори. Так, при змішуванні спектральних кольорів не можна отримати таких кольорів, як брунатний, хакі, маслиновий. Вважають, що неспектральні кольори виникають тільки ири змішуванні пігментів, зокрема їх можна отримати шляхом додавання до спектральних кольорів "чорного кольору". Це здійснюється не лише за допомогою пігментів, а й приладів: колірної телевізійної установки, багатоканального колірного стимулятора.
Так, жовтогаряча пляма в оточенні білого фону буде жовтогарячою лише за умови, що яскравість самої плями не менша яскравості фону. Якщо ж зробити яскравість фону більшою, ніж яскравість плями, тобто змістити точку в колірному просторі по осі У/кііе — Віаск відносно нульового рівня, то отримаємо насичений брунатний колір.
Отже, це доводить, що "чорна" половина колірного простору реально існує. Власне кажучи, на цьому ґрунтується кольорове телебачення. Люмінофори телевізійного екрана, як і всі світлові джерела, створюють тільки спектральні випромінювання. Проте внаслідок усереднення освітленості на екрані телевізора сітківкою ока ті ділянки екрана, освітленість яких вища за середній рівень, сприймаються забарвленими у спектральні кольори, а ті ділянки, освітленість яких нижча за середній рівень, сприймаються забарвленими у неспектральні кольори.
Протягом першого року житгя у дитини спостерігається повна колірна сліпота. Відчуття червоного з'являється у дівчаток у 14, зеленого — у 16 і синього — у 18 місяців. У хлопчиків цей процес відбувається на 2 місяці пізніше. Діти віком 2,5 — З років досить добре визначають червоний