

1.Обмен веществ и энергии в клетке. Классификация организмов по способу питания: автотрофы (фототрофы и хемотрофы), гетеротрофы, миксотрофы. Обмен веществ или метаболизм -процесс превращения в-в в живой материи, лежащий в основе жизни, совокупность химических реакций в организме, которые обеспечивают его веществами и энергией, необходимыми для жизнедеятельности. Анаболизм (ассимиляция) + Катаболизм (диссимиляция) = Метаболизм. Обмен вещ-в и превращение энергии - основа жизнедеятельности клеток. Обмен вещ-в - последовательное потребление, превращение, использование, накопление и потеря вещ-в и энергии в живых организмах в процессе жизнедеятельности, что позволяет им самосохраняться, развиваться и самовоспроизводиться.В процессе ассимиляции (анаболизма) происходит синтез из простых веществ более сложных, крупных молекул, с поглощением энергии. В процессе диссимиляции (катаболизма) происходит расщепление, сложных вещ-в до простых с участием ферментов, с освобождением энергии. Обмен вещ-в можно рассматривать как общий и по разным группам вещ-в - белковый, жировой, углеводный, водно-солевой. У каждого из них имеются свои системы регулирования. Ассимиляция и диссимиляция - две стороны единого процесса обмена веществ. Анаболизм - пластический обмен, ассимиляция -происходит в три этапа: 1).синтез промежуточных соединений из низкомолекулярных вещ-в (органич. кислот, альдегидов); 2)синтез" строительных блоков" из промежуточных соединений (аминокислот, жирных кислот, моносахаридов); 3) синтез из "строительных блоков" макромолекул белков, нуклеиновых кислот, полисахаридов, жиров. Идёт с поглощением энергии, участием ферментов. Катаболизм- энергетический обмен, диссимиляция или процесс расщепления сложных орг. вещ-в идёт в три этапа: 1.Подготовительный – сложные орг. вещ-ва ферментативно превращаются в простые и выделяется немного энергии. 2-й этап – бескислородный – дальнейший распад веществ. Напр. гликолиз глюкозы С6Н12О6 – 2С3Н6О3 - свободная энергия. 3-й этап – кислородный, окончательного расщепления орг. вещ-в до простых (СО2 и Н2О) + макс.энер По способу питания, а значит, по способам потребления энергии организмы подразделяются на три группы. А).Автотрофные организмы (автотрофный тип питания) – способны синтезировать органические вещества из неорганических. Автотрофные организмы фотосинтезирующие (фотоавтотрофы), к которым относятся зеленые растения и фотосинтезирующие бактерии, при создании органических соединений непосредственно используют лучистую энергию Солнца – единственного источника энергии для живой природы Земли. Все остальные живые существа используют энергию, заключенную в химических связях. Автотрофные хемосинтезирующие организмы (хемоавтотрофы), к которым относятся некоторые бактерии, для синтеза органических соединений применяют энергию, выделяющуюся при окислении неорганических соединений (сероводорода, аммиака, железа и др) Б).Гетеротрофные (гр.heteros –иной, другой) организмы (животные, грибы, не зеленые растения, большинство бактерий) не способны самостоятельно синтезировать органические вещества из неорганических, они используют энергию химических связей готовых органических соединений (гетеротрофный тип питания). Гетеротрофные организмы, в свою очередь, подразделяются на сапрофитов (гр.sapros-гнилой +phagos –пожирающий) и паразитов. Сапротрофы, или сапротрофные организмы, питаются органическими веществами мертвых тел (большинство видов животных, бактерий и грибов). Паразиты, или паразитические организмы (болезнетворные бактерии, паразитические растения, животные, грибы), потребляют органические вещества живых организмов В).Миксотрофные (греч.mixis-смешение +trophe –питание) организмы, например, эвглена зеленая, насекомоядные растения (миксотрофный тип питания) могут питаться и как автотрофы, и как гетеротрофы. Общая схема обмена веществ и энергии: 1 - пищеварение; 2 - катаболизм; 3 - анаболизм; 4 - распад структурно-функциональных компонентов клеток; 5 - экзергонические реакции – (катаболические) – это реакции распада крупных молекул на мелкие с выделением энергии; 6,7 – эндергонические - анаболические (эндотермические) - реакции синтеза крупных молекул с затратой энергии; 8 - выведение из организма.
2.Автотрофное питание: фотосинтез и хемосинтез. Фотосинтез— процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (напр. хлорофилла у растений). Возникновение фотосинтеза и кислородного дыхания — важнейшие ароморфозы архейской эры. Общий вид фотосинтеза - 6СО2 +6 Н2О + С.Э.=С6Н12О6 + О2.. Автотрофные фотосинтезирующие организмы (фотоавтотрофы), к ним относятся зеленые растения и фотосинтезирующие бактерии, при создании органических соединений используют энергию Солнца. Все остальные живые существа используют энергию, химических связей. Автотрофные хемосинтезирующие организмы (хемоавтотрофы), некоторые бактерии, для синтеза органических соединений применяют энергию, выделяющуюся при окислении неорганических соединений (сероводорода, аммиака, железа и др.) Все животные существуют за счет орган. в-ва, созданного автотрофами, и аккумулированной в этом вещ-ве солнечной энергии. Возникновение фотосинтеза (3—3,5 млрд. лет назад) обусловило разделение органического мира на автотрофный и гетеротрофный. Сине-зеленые водоросли были первыми фотосинтезирующими организмами. С появлением фотосинтеза появился надежный источник органических веществ на Земле, а в атмосферу планеты стал выделяться кислород. Существует два различных вида фотосинтеза: В первом случае одним из источников ионов водорода и электронов служит вода, расщепляющаяся под действием фотонов света, а основным акцептором атомов водорода и источником углерода – СО2. Кроме того, одним из конечных продуктов является кислород, выделяющийся в атмосферу. Такой тип фотосинтеза называется кислородным, или оксигенным. Характерен для всех зеленых растений (эукариот) и цианобактерий (прокариот). При использовании в фотосинтезе только CO2 и H2O образуются углеводы. Но в процессе фотосинтеза растения образуют не только углеводы, но и содержащие азот и серу аминокислоты, белки, а также пигменты и др. соединения. Акцепторами атомов водорода в данном случае (наряду с CO2) и источниками азота и серы при этом служат нитраты (NO3-) и сульфаты(SO4-). Во втором случае вода не используется, а донором водорода являются неорганические (сероводород, тиосульфат, газообразный водород) и органические соединения (молочную кислоту, изопропиловый спирт). Источником углерода в большинстве случаев является также CO2, но наряду с этим и некоторые органические соединения (например, ацетат). В результате такого фотосинтеза кислород не образуется и не выделяется. Такой тип фотосинтеза называется бескислородным, или аноксигенным.( Встречается у зеленых и пурпурных серных и несерных бактерий). В обоих случаях фотосинтез идет на сложноорганизованных мембранах, где для синтеза АТФ используется энергия трансмембранного электрохимического градиента ионов водорода (DН+) - Кислородный, или оксигенный фотосинтез. Фотосинтез идет в 2 стадии. Первая получила название световой стадии фотосинтеза (фотофизический и фотохимический этапы); вторая – темновой стадии фотосинтеза (химический этап).
3.Световая
фаза фотосинтеза. Фотосинтетическое
фосфорилирование: циклическое и
нециклическое.
В
световой фазе два этапа. На первом
этапе – фотофизическом -
происходит поглощение квантов света
пигментами, их переход в возбуждённое
состояние и передача энергии к другим
молекулам фотосистемы. На втором
этапе –фотохимическом - происходит
разделение зарядов в реакционном центре,
перенос электронов по фотосинтетической
электрон-транспортной цепи, что
заканчивается синтезом АТФ и НАДФ.Н2.
Вместе первые два этапа называют
светозависимой стадией фотосинтеза.
Центральным событием фотосинтеза
(преобразование световой энергии в
химическую) является реакционный центр.
Когда молекулы вспомогательных пигментов
передают энергию на одну молекулу
главного пигмента. Энергия света в нём
и используется для осуществления
химической реакции. Посредником во
взаимодействии этих соединений с
квантами служит хлорофилл, включенный
в структуру хлоропласта или хроматофора.
Каротиноиды поглощают свет с длиной
волны 280–550 нм (это зеленая, синяя,
фиолетовая, ультрафиолетовая области
спектра).. Чем больше в молекуле сопряженных
двойных связей, тем больше длина волны
поглощаемого света.. Соответственно
меняется и окраска пигмента. Первая
функция каротиноидов - светособирающая
- поглощают сине-зеленый свет и передают
его энергию хлорофиллам.. Эта функция
каротиноидов особенно важна для
водорослей, поскольку сине-зеленый свет
проникает в толщу воды гораздо глубже,
чем красный.. Вторая функция каротиноидов
в хлоропластах — светозащитная - защищают
хлорофилл фотосистем от светового
«перевозбуждения». Третья функция
каротиноидов — структурная - служат
необходимыми структурными элементами
фотосистем. Пигменты фотосинтеза
растений располагаются на внутренней
стороне мембраны тилакоидов.. Они
организованы в фотосистемы – целые
цеха по улавливанию света – каждая
система содержит 250-400 молекул разных
пигментов и представляет собой
обособленную структуру мембраны. В
реакционном центре происходит основной
акт фотосинтеза – разделение зарядов
с последующим образованием первичного
окислителя и первичного восстановителя.
Существуют два типа центров, один из
которых включен в пигментную фотосистему
I (ФС I или ФС1), а другой – в фотосистему
II (ФС II или ФС2). В фотосистеме I
реакционный центр представлен хлорофиллом
а с максимумом поглощения света при
длине волны 700 нм. В фотосистеме
II – хлорофиллом а с максимумом
поглощения света при длине волны 680 нм.
Комплексы ФС2 локализованы в основном
в уплотненных областях тилакоидов гран,
а комплексы ФС1 находятся преимущественно
в стромальных участках тилакоида и в
неуплотненных областях тилакоидов
гран. Фотосистема II — сложный
пигмент-белковый комплекс.
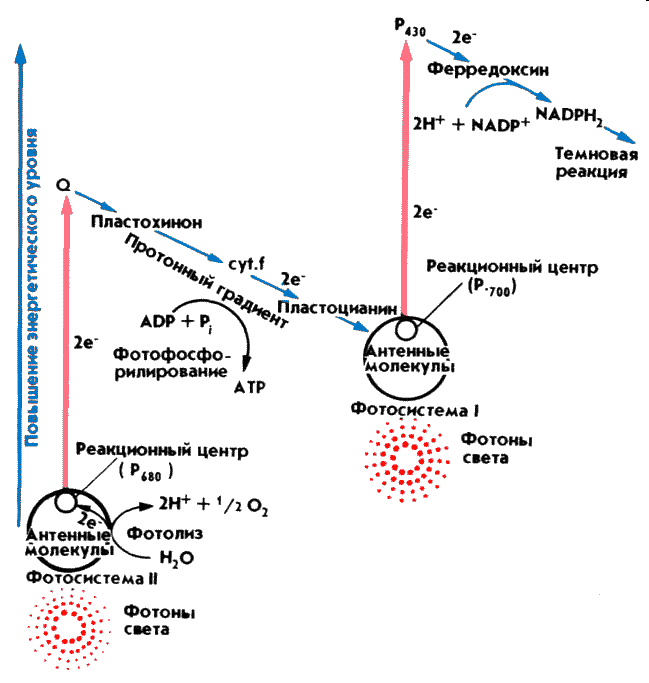
Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах. Р680 в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула Р680 становится сильным восстановителем– первичный донор электронов. После потери электрона хлорофилл переходит в основное состояние, и этим заканчивается первый этап фотосинтеза, называемый фотофизическим. Фотохимическая фаза фотосинтеза с участием ФС II. Первичным акцептором является молекула феофитина. Далее электрон переносится на хиноны и далее на пластохиноны (PQ), транспортируемый в восстановленной форме к суt.f комплексу. Одна молекула пластохинона является двухэлектронным акцептором и может быть как связана с ФС2, так и свободно отделяться от ФС2. Получив в результате двукратного срабатывания фотосистемы II два электрона и присоединив два протона из стромального пространства, дважды восстановленный пластохинон (PQH2) отделяется от реакционного центра ФС2 и начинает диффундировать внутри липидной мембраны тилакоида. Место связывания оказывается свободным, и к нему может присоединиться новая молекула PQ.
Фотолиз воды .Заполнение электронной вакансии в молекуле H680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре 4 иона марганца. Комплекс находится со стороны внутритилакоидного пространства и образующие в результате фотолиза воды свободные электроны перемещаются в молекулу хлорофилла, где занимают место ранее выбитых фотонами света электронов, а полученные протоны выбрасываются во внутритилакоидное пространство (люмен). 2Н2О 4Н+ + 4е- + О2. Таким образом, суммарный результат работы ФС- II — это окисление 4 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране. Фотосистема I — сложный пигмент-белковый комплекс. Цепь электронного транспорта фотосистемы I функционирует следующим образом: после возбуждения Р700 квантом света и перехода в возбужденное состояние электрон очень быстро переносится на Р. После чего электрон достигает ферредоксина, который покидает сайт связывания и переносит электрон дальше по цепи к ферменту ферредоксин-НАДФ-редуктазе, который связан с внешней стороной мембраны тилакоида и катализирует восстановление НАДФ+ -следующего акцептора электронов до НАДФН в соответствии с реакцией: 2Fdred + НАДФ+ + H+ => 2Fdox + НАДФН В восстановленной форме молекулы NADPH принимают участие в цикле фиксации углерода (цикле Кальвина).
Цитохромный комплекс b6–f, в состав которого входят цитохром f и два цитохрома b6, а также медьсодержащий белок пластоцианин (Pc). Он является связующим звеном электронтранспортной цепи фотосинтеза. Цитохромный комплекс - занимает положение между ФС II и ФС I, контролируя общую скорость транспорта электронов, и обеспечивая баланс между притоком электронов (от ФС II в случае нециклического и от ферредоксина в случае циклического транспорта) и оттоком электронов из ФС I. Важнейшей функцией цитохромного комплекса является сопряжение транспорта электронов по электронтранспортной цепи с формированием электрохимического потенциала протонов (ΔμH+) на мембране тилакоида, который используется для синтеза АТФ.
Фотофосфорилирование. Окисление РQH2 цитохромным комплексом b6f осуществляется в реакциях Q-цикла. Благодаря работе Q-цикла происходит направленный перенос протонов через мембрану. В результате Q-цикла, а также фотолиза фоды протоны (Н+) накапливаются внутри мембраны тилакоидов, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны. При достижении между двумя сторонами мембраны критической разности потенциалов (200 мВ) протоны устремляются по протонному каналу в ферменте АТФ-синтетазе наружу. На выходе из канала создается высокий уровень энергии, которая идет на синтез АТФ. Для синтеза одной молекулы АТФ необходимо, чтобы три иона водорода вышли из тилакоида в строму хлоропласта. Результат световой фазы фотосинтеза при нециклическом фотофосфорилировании (а это основной вариант) можно записать в виде реакции: 2NADP+ 2АДФ + 2Ф- + 2H2O + 4 hv(квант света)= 2NADPH + 2АТФ + O2, т.е в результате энергия света, заключенная в фотонах идет на синтез АТФ и НАФН (3 АТФ) Фотосистема I может работать и автономно. Происходит это при замедлении или прекращении окисления НАДФН(нехватке НАД+) и избытке света. При этом используется обходной путь переноса электронов от возбужденного реакционного центра – а именно та же цепь переноса электронов, которая ведет из фотосистемы II. Электроны проходят по ней, вызывая. сопряженный транспорт протонов из внешней среды тилакоида во внутреннюю, чем усиливается протонный градиент, и возвращаются обратно к реакционному центру фотосистемы I. Таким образом, здесь нет никаких окислительно-восстановительных реакций (кислород при этом не выделяется и НАДФ не восстанавливается). Это называется циклическим фотофосфорилированием. Оно может идти параллельно с нециклическим. Кроме того, используется некоторыми фотосинтетическими бактериями, которые в процессе фотосинтеза не выделяют кислорода.Псевдоциклическое фотофосфорилирование •е-от воды переносится не на НАДФ+, а на О2 с образованием, в конечном счёте, воды. •Включается, когда мало НАДФ+. •Идёт в ФС1 и ФС2. •Вредно – образуются активные формы кислорода
4. Фотодыхание. Темновая стадия фотосинтеза (в строме) : с3-, с4- и сам-типы фотосинтеза. *
Фотодыхание – это светозависимый процесс, т.к. рибулозобисфосфат – один из продуктов цикла Кальвина – образуется только тогда, когда идёт фотосинтез. (Назначение фотодыхания – вернуть в цикл хотя бы часть углерода из гликолата.) Цикл Кальвина (С3-путь) или восстановительный пентозофосфатный цикл -главный путь ассимиляции СО2, , представляет собой циклический процесс, в который вводится СО2 и из которого выходит углевод. Процесс можно разделить на три фазы: первая фаза - непосредственная фиксация СО2 – карбоксилирование: СО2, связываясь с рибулозобифосфатом, образует две молекулы фосфоглицерата. Эту реакцию катализирует рибулозобифосфат-карбоксилаза. Вторая фаза - восстановление. Фосфоглицерат при участии NADPН (восстановитель) и АТР (донор энергии) восстанавливается до 3-фосфоглицеральдегида (ФГА). Третья фаза – образование продуктов фотосинтеза. Четвертая стадия- восстановление первоначальных реагентов. Для того,чтобы растение могло акцептировать новую молекулу СО2, необходимо иметь РДФ, основной акцептор СО2. РДФ образуется из ФГА в результате цепи реакций, в процессе которых образуются пяти- и семиуглеродные сахара. (Из 12 образовавшихся молекул ФГА только две идут на образование продуктов фотосинтеза.) С окончанием этой фазы цикл замыкается. Ферменты цикла находятся в строме хлоропласта, а рибулозобифосфат-карбоксилаза - на наружной стороне тилакоидных мембран. Суммарное уравнение синтеза глюкозы в цикле Кальвина: 6CO2 + 12НAДФH + 18ATФ → C6H12O6 + 12НAДФ + 18AДФ + 6H2O Фотосинтез по С3-пути всегда сопровождается фотодыханием – процессом потребления на свету кислорода и выделения СО2. фотодыхание – довольно затратный процесс, т.к. он не сопровождается синтезом АТФ, а, наоборот забирает энергию для восстановления кислорода. Более того, 50% углерода, используемого при фотосинтезе у С3-растений, восстанавливается до СО2. Образование продуктов фотосинтеза. Обычно продуктом фотосинтеза называют сахар. На самом деле продуктами фотосинтеза можно считать и др. вещества. Молекула ФГА используется растением в цикле Кальвина в нескольких направлениях. Во-первых, ФГА является основой для синтеза сахара. Во-вторых, ФГА может быть использована для синтеза аминокислот. Синтез аминокислот происходит интенсивно при недостатке НАДФН, в результате чего из фосфоглицериновой кислоты образуется не ФГА, а пировиноградная кислота, которая является исходным соединением для синтеза аминокислот и одним из ключевых веществ цикла Кребса. В-третьих, ФГА дает начало циклу превращений некоторых промежуточных продуктов в РДФ, который служит акцептором СО2. Наряду с углеводами и аминокислотами из промежуточных продуктов цикла Кальвина могут образовываться липиды и др. продукты. Для фиксации 1 молекулы СО2 нужны 2 молекулы НАДФН и 3 молекулыАТФ. Запаса энергии (53,2*104Дж=-132ккал) этих молекул достаточно, чтобы восстановить одну молекулу СО2 до глюкозы. Энергия одного моля кванта красного света с длиной волны 680 нм равна 17,6*104 Дж, соответственно, для фиксации 1 молекулы СО2 нужно по крайней мере 3 кванта света. В действительности нужно не менее 8 квантов (4 кванта, по одному на перенос каждого из четырех электронов необходимых для выделения молекулы О2, умножаются на 2 – число световых реакций), чтобы одновременно восстановить НАДФ+ и синтезировать АТФ в нужных количествах. При фиксации СО2 в процессе фотосинтеза используется лишь около 30% энергии света (потребляется 8-10 квантов там, где должно было хватить энергии 2,7 кванта). Средняя эффективность использования фотосинтетически активного солнеч. света растениями не превышает 1%.
Для С4 растений характерна особая морфология листа и они содержат хлоропласты двух различных типов. Хлоропласты находятся в клетках листа, расположенных вокруг сосудистого пучка двумя концентрическими слоями: внутренний слой –клетки обкладки сосудистого пучка, хлоропласты довольно крупные, не имеют, как правило гран и содержат много зерен крахмала. внешний – клетки мезофилла, хлоропласты мезофилла довольно мелкие расположены в клетке беспорядочно, содержат стопки гран и небольшое количество зерен крахмала. У С4-растений фиксация СО2 может осуществляться двумя путями:обычный цикл Кальвина, функционирующий в клетках обкладки. Первичный продукт фиксации СО2 – фосфоглицериновая кислота – С3.В хлоропластах мезофилла фиксация углекислоты происходит в результате присоединения СО2 к фосфоенолпирувату, приводящему к образованию кислот четырьмя атомами углерода – оксалоацетата и малат – это «С4-путь» фиксации СО2.
САМ-типы: 1. Их устьица обычно открыты ночью (т.е. в темноте) и закрыты в течение дня. 2. Фиксация углекислого газа происходит в темное время суток с образованием кислот, содержащих четыре атома углерода – оксалоацетата и малата. 3. Малат запасается в больших вакуолях, которые характерны для клеток МОКТ-растений.4. В светлое время суток малат переносится в цитоплазму и там декарбоксилируется, образуя пируват и СО2, которая поступает в цикл Кальвина, где превращается в сахарозу или запасной углевод глюкан. 5. В темновой период суток часть запасенного глюкана распадается с образованием молекул-акцепторов для темновой фиксации углекислого газа