

Модель Вольтерра и ее обобщения
Перейдем к описанию конкретных моделей. Начнем с классической модели Вольтерра и ее обобщений. В основу своих работ Вольтерра положил следующие простые гипотезы:
Пища либо имеется в неограниченном количестве, либо ее поступление с течением времени лимитировано.
Существует естественная смертность особей каждого вида.
Хищники поедают жертв, причем количество съеденных жертв пропорционально вероятности встречи хищника и жертвы, т.е. произведению количества хищников на количество жертв.
Если имеется несколько видов потребляемой пищи, имеющейся в ограниченном количестве, то доля потребляемой в единицу времени пищи пропорционально количеству особей.
Если вид питается пищей, количество которой неограниченно, то его прирост пропорционален численности.
Если пища имеется в ограниченном количестве, то прирост численности вида пропорционален количеству съеденной пищи.
Гипотезы 1)-6) в сущности аналогичны представлениям классической химической кинетики и закону действующих масс.
Для двух видов, численность которых обозначается как и , в соответствии с гипотезами Вольтерра 1)-6) можно записать следующую систему уравнений:
![]() (
4.24)
(
4.24)
Если
положить
![]() ,
т.е. взаимодействие между видами
отсутствует, то мы получаем два
логистических уравнения, решения которых
хорошо известны. Если хотя бы один их
коэффициентов
,
т.е. взаимодействие между видами
отсутствует, то мы получаем два
логистических уравнения, решения которых
хорошо известны. Если хотя бы один их
коэффициентов
![]() и
и
![]() отличен от нуля, то виды начинают
взаимодействовать.
отличен от нуля, то виды начинают
взаимодействовать.
Рассматривая различные комбинации значений коэффициентов , , выделим три важнейших класса:
 – симбиоз;
– симбиоз;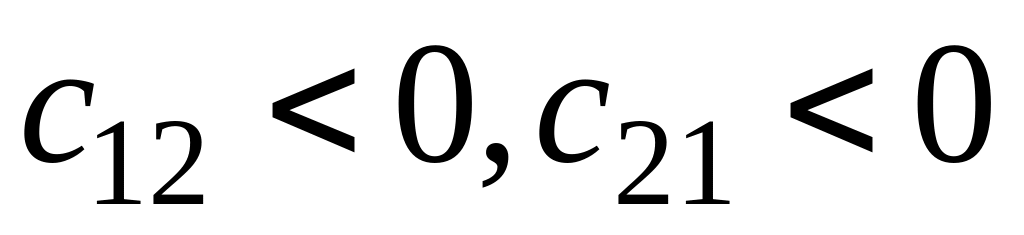 – конкуренция;
– конкуренция; – хищник–жертва.
– хищник–жертва.
Наибольший интерес представляют для нас два последних случая: конкуренция и хищник–жертва.
Рассмотрим стационарные состояния системы (4.24), которые определяются из следующих уравнений:
![]() (
4.25)
(
4.25)
Система (4.25) имеет четыре решения:
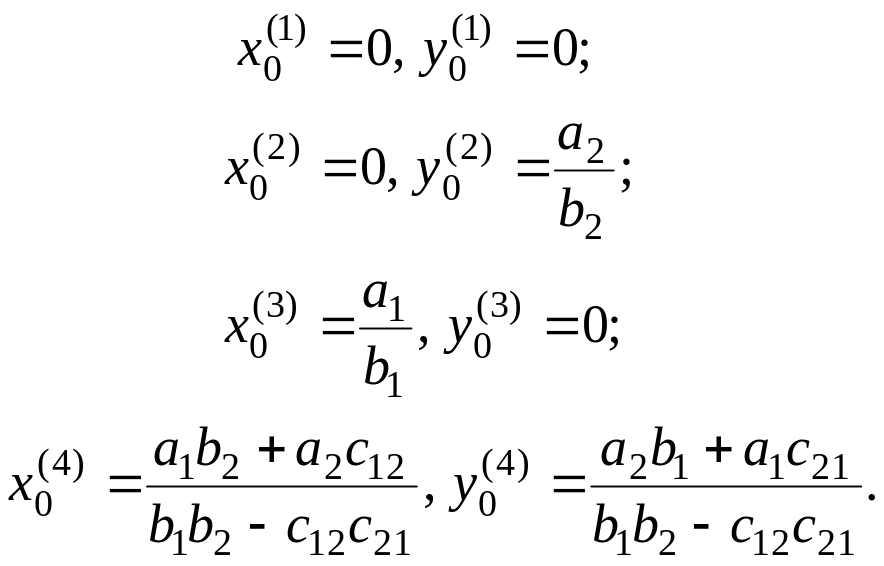 (
4.26)
(
4.26)
Перейдем теперь к модели конкуренции ( ) и исследуем стационарные состояния:
Стационарное состояние (0,0) всегда неустойчивый узел.
Стационарное состояние
 является седлом при
является седлом при
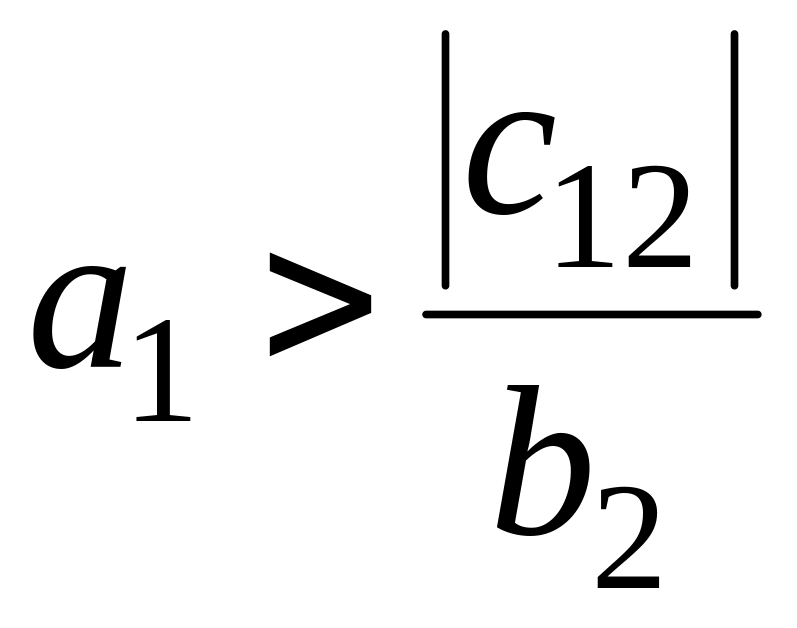 и устойчивым узлом в противном случае.
Это означает, что вид вымирает, если
его собственная скорость роста меньше
некоторой критической величины.
и устойчивым узлом в противном случае.
Это означает, что вид вымирает, если
его собственная скорость роста меньше
некоторой критической величины.Стационарное состояние
 аналогично (2).
аналогично (2).Это стационарное состояние характеризует сосуществование двух конкурирующих видов и в ряде случаев представляет собой устойчивый узел.
Сделав линейную замену переменных в системе (4.24), можно привести ее к виду, содержащего всего три параметра:
![]() (
4.27)
(
4.27)
где стационарные точки определяются как
 .
( 4.28)
.
( 4.28)
Проанализируем возможные типы фазовых портретов. Для этого рассмотрим так называемые главные изоклины системы. Напомним определения:
1)
Изоклиной
на фазовой плоскости
![]() называется кривая, такая, что в каждой
ее точке вектор скорости
называется кривая, такая, что в каждой
ее точке вектор скорости
![]() принимает некоторое постоянное значение.
принимает некоторое постоянное значение.
2)
Главными
изоклинами
называются изоклины, для которых либо
![]() ,
либо
,
либо
![]() .
.
Очевидно, что пересечение главных изоклин дает стационарные точки системы.
В данном случае имеется всего 4 главных изоклин, представляющих собой прямые линии.
Изоклины,
на которых
![]() –
–
![]() ;
изоклины, на которых
;
изоклины, на которых
![]() –
–
![]() .
.
Заметим,
что сами оси
![]() и
и
![]() являются изоклинами.
являются изоклинами.
В первой четверти декартовой плоскости мы имеем четыре различных типа фазовых портретов, показанных на рис. 4.12.
(а)
![]() .
Здесь внутри положительного квадранта
существует точка пересечения изоклин,
которой соответствует точка сосуществования
двух видов. Векторное поле скоростей
показывает, что это стационарное
состояние устойчиво (устойчивый узел).
.
Здесь внутри положительного квадранта
существует точка пересечения изоклин,
которой соответствует точка сосуществования
двух видов. Векторное поле скоростей
показывает, что это стационарное
состояние устойчиво (устойчивый узел).
|
Рис. 4.12. Качественно различные типы фазовых портретов в первом квадранте. Стационарные точки показаны красными кружочками, а изоклины (кроме осей координат) – голубыми линиями
|
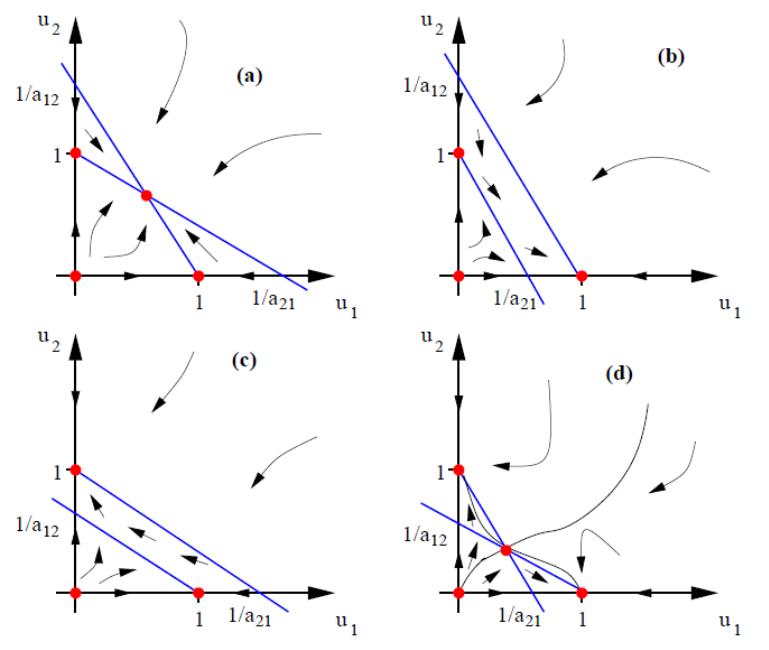
(b)
![]() .
Здесь стационарные состояния расположены
на границах положительного квадранта,
т.е. сосуществование видов невозможно.
.
Здесь стационарные состояния расположены
на границах положительного квадранта,
т.е. сосуществование видов невозможно.
(c)
![]() .
Случай полностью аналогичен случаю
(b).
.
Случай полностью аналогичен случаю
(b).
(d)
![]() .
Как и в случае (а), существует стационарное
состояние, соответствующее сосуществованию
видов, однако, в данном случае оно
неустойчивое (седло) и устойчивыми
являются граничные стационарные
состояния (0,1) и (1,0). Это означает, что в
системе существует множественность
стационарных состояний, т.е. в зависимости
от начальных условий система стремится
либо к стационарному состоянию (1,0), либо
к стационарному состоянию (0,1).
.
Как и в случае (а), существует стационарное
состояние, соответствующее сосуществованию
видов, однако, в данном случае оно
неустойчивое (седло) и устойчивыми
являются граничные стационарные
состояния (0,1) и (1,0). Это означает, что в
системе существует множественность
стационарных состояний, т.е. в зависимости
от начальных условий система стремится
либо к стационарному состоянию (1,0), либо
к стационарному состоянию (0,1).
Основной
результат, полученный нами, сводится с
математической точки зрения к следующему
утверждению: Устойчивое стационарное
состояние внутри первого квадранта
существует в случае
![]() ,
т.е. когда взаимодействие между двумя
видами достаточно слабо.
,
т.е. когда взаимодействие между двумя
видами достаточно слабо.
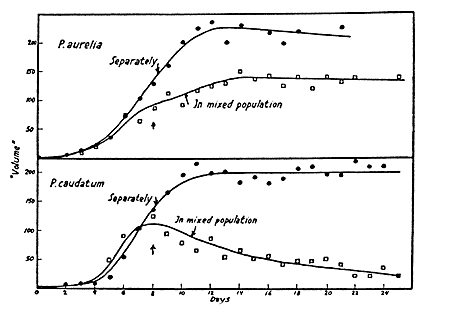
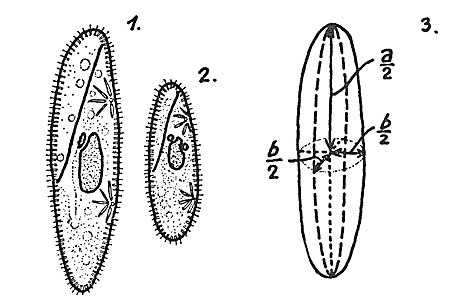
На языке биологии мы получаем следующее фундаментальное утверждение: Закон конкурентного исключения (Г. Гаузе) – в одной экологической нише может существовать только один вид. На рис. 4.13 приведены результаты эксперимента Гаузе для двух видов инфузорий-туфелек Paramecium. Туфельки –это крупные одноклеточные эукариоты, длина которых в зависимости от вида (их несколько сотен) колеблется от 50 до 350 микрон. Paramecium Aurelia и Paramecium Caudatum, использованные в экспериментах Гаузе, показаны на рис. 4.14. На рис. 4.15 показаны относительные размеры этих двух видов.
Рассмотрим
теперь случай разных знаков коэффициентов
![]() ,
,
![]() .
Это одна из простейших моделей
«хищник-жертва» или модель Лотки-Вольтерра
с логистической поправкой. Ее можно
исследовать тем же самым способом, что
и проанализированная подробно модель
конкуренции двух видов. Мы не будем
этого делать, а приведем лишь самый
главный результат. В этой модели ни при
каких значениях параметров невозможны
устойчивые (незатухающие) колебания
численности обоих видов. Стационарное
состояние, соответствующее сосуществованию
хищника и жертвы, может быть либо
устойчивым, либо неустойчивым фокусом.
Это означает, что мы будем наблюдать
либо затухающие колебания, либо, наоборот,
растущие.
.
Это одна из простейших моделей
«хищник-жертва» или модель Лотки-Вольтерра
с логистической поправкой. Ее можно
исследовать тем же самым способом, что
и проанализированная подробно модель
конкуренции двух видов. Мы не будем
этого делать, а приведем лишь самый
главный результат. В этой модели ни при
каких значениях параметров невозможны
устойчивые (незатухающие) колебания
численности обоих видов. Стационарное
состояние, соответствующее сосуществованию
хищника и жертвы, может быть либо
устойчивым, либо неустойчивым фокусом.
Это означает, что мы будем наблюдать
либо затухающие колебания, либо, наоборот,
растущие.
|
Рис. 4.13. Результаты эксперимента Гаузе для двух видов инфузорий-туфелек Paramecium: сверху рисунка P. aurelia, снизу – P. caudatum. По горизонтальной оси отложены дни, по вертикальной оси – объем популяций туфелек. Заполненные (незаполненные) кружки показывают экспериментальные данные для популяций без конкуренции (с конкуренцией).
|
|
|
а) |
б) |
Рис. 4.14. Paramecium Aurelia (а) и Paramecium Caudatum (б), использованные в экспериментах Гаузе |
|
|
Рис. 4.15. Относительные размеры двух видов: 1. P. caudatum; 2. P. aurelia
|
Рассмотрим этот вопрос подробнее. Проанализируем оригинальную модель Лотки-Вольтерра, которая является упрощенным вариантом модели, описанной выше. Отличие заключается в том, что вместо логистического вида правых частей уравнения в отсутствии межвидового взаимодействия (то, что использовалось нами) в исходных уравнениях
|
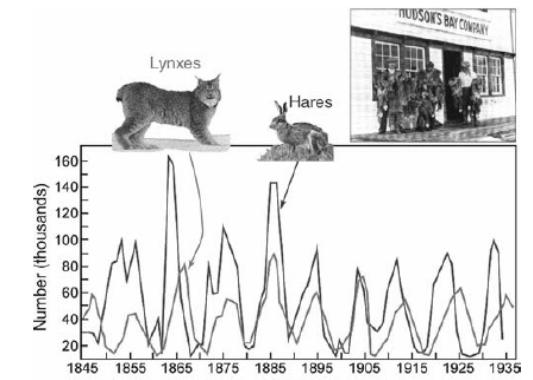
Рис. 4.16. Количество шкурок рысей и зайцев, сданных на приемные пункты компании «Hudson’s Bay Company» (Канада). По горизонтальной оси отложены годы, по вертикальной оси – количество шкурок в тысячах штук |
Лотки-Вольтерра правые части имели «мальтузианский» вид – неограниченное экспоненциальное размножение жертв и такая же экспоненциальная смерть хищников. Запишем эти уравнения, сохранив наши обозначения, в следующем виде:
![]() (
4.29)
(
4.29)
Здесь
![]() и
и
![]() .
Для простоты обозначения положив
.
Для простоты обозначения положив
![]() ,
,
![]() ,
из (4.29) получим
,
из (4.29) получим
![]() (
4.30)
(
4.30)
Проведем,
так же, как и ранее, исследование
стационарных состояний. Теперь их только
два: (0,0) и
![]() .
Покажем, что второе стационарное
состояние, соответствующее сосуществованию
популяций, является центром. Действительно,
линеаризованная система в этой точке
принимает следующий вид:
.
Покажем, что второе стационарное
состояние, соответствующее сосуществованию
популяций, является центром. Действительно,
линеаризованная система в этой точке
принимает следующий вид:
![]() (
4.31)
(
4.31)
Запишем характеристическое уравнение:
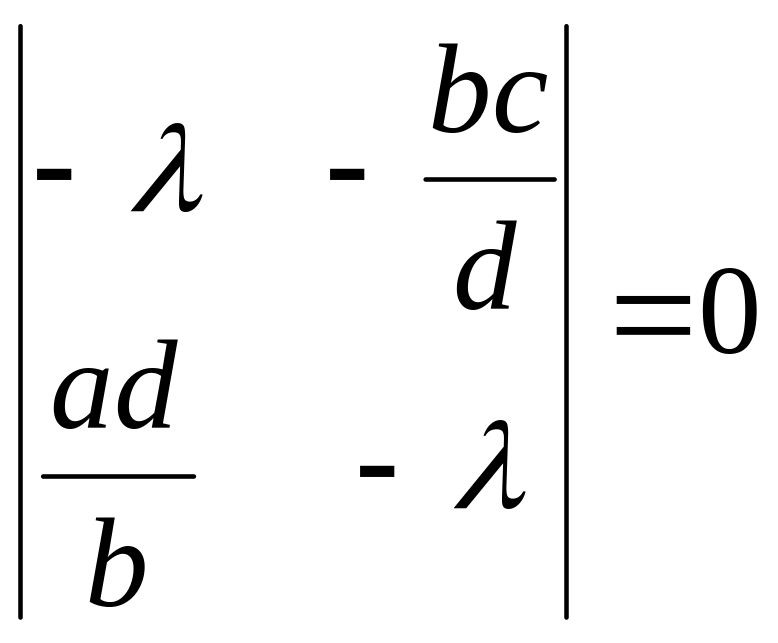 .
( 4.32)
.
( 4.32)
Корни
уравнения (4.32)
![]() .
Следовательно, стационарное состояние
является центром. Кинетические кривые
системы (4.30) показаны на рис. 4.17.
.
Следовательно, стационарное состояние
является центром. Кинетические кривые
системы (4.30) показаны на рис. 4.17.
|
Рис. 4.17. Кинетические кривые системы (4.30). По горизонтальной оси отложено время (в усл.ед.), по вертикальной оси – объем популяций (в усл.ед.) |

Видно, что численность обеих популяций меняется периодически, т.е. имеют место незатухающие колебания. Казалось бы, все хорошо и мы имеем модель, достаточно адекватно отображающую реальную действительность. Однако, к сожалению, стационарная точка, вокруг которой происходят колебания, является центром. А это – негрубая стационарная точка, т.е. она неустойчива относительно сколь угодно малых изменений правой части уравнения. Именно этот эффект мы видели в более сложной модели с логистическими добавками, в которой центр превращается в устойчивый или неустойчивый фокус. Таким образом, на самом деле исходная модель Лотки-Вольтерра не описывает незатухающие колебания, имеющие место в реальных системах. По существу мы приходим к выводу, что для описания незатухающих колебаний численности популяций необходимо построение моделей более сложных, чем рассмотренные выше.
Рассмотрим соответствующий пример. Это так называемая модель Розенцвейга-Макартура. В общем виде она записывается как
![]() (
4.33)
(
4.33)
где
– скорость изменения численности жертв
в отсутствии хищников,
![]() – скорость уменьшения количества жертв
из-за наличия хищников,
– коэффициент смертности хищников (в
«мальтузианском» виде),
– коэффициент эффективности усвоения
жертв хищниками.
– скорость уменьшения количества жертв
из-за наличия хищников,
– коэффициент смертности хищников (в
«мальтузианском» виде),
– коэффициент эффективности усвоения
жертв хищниками.
Вид функций и естественно меняется от модели к модели в зависимости от конкретной биологической системы.
Приведем одну из таких моделей, предложенной Макартуром, которая описывает взаимодействие двух видов насекомых:
![]() (
4.34)
(
4.34)
В
первом уравнении описываются следующие
процессы: насекомые вида
поедают личинок вида
(член
![]() ),
но взрослые особи вида
поедают личинок вида
при условии высокой численности видов
или
,
или обоих видов (члены
),
но взрослые особи вида
поедают личинок вида
при условии высокой численности видов
или
,
или обоих видов (члены
![]() ).
).
Во
втором уравнении член
![]() отражает скорость прироста вида
;
отражает скорость прироста вида
;
![]() – логистическая добавка;
– логистическая добавка;
![]() – поедание личинок вида
насекомыми вида
;
– поедание личинок вида
насекомыми вида
;
![]() – увеличение популяции вида
за счет поедания личинок вида
.
– увеличение популяции вида
за счет поедания личинок вида
.
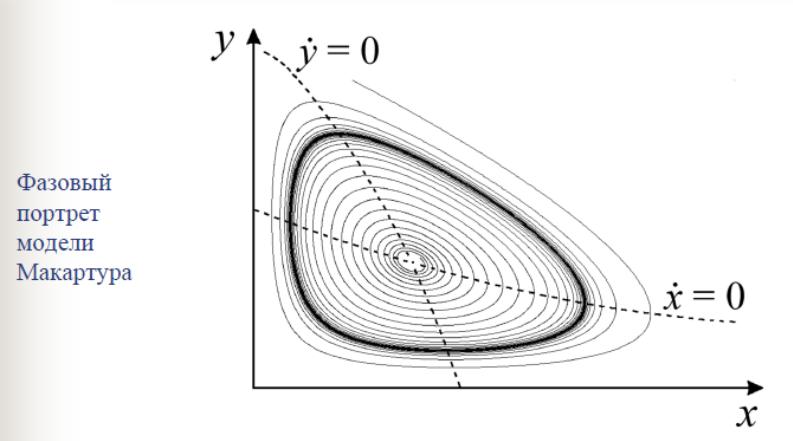
|
Рис. 4.18. Фазовый портрет модели Макартура
|
Популяционная динамика, начиная с работ Вольтерры – традиционный объект математической биологии. Простейшим объектом моделирования являются микробные сообщества, наподобие рассмотренным нами сообществ туфелек. А сложнейшим и уникальным объектом является вся биосфера Земли. На этом мы закончим обзор моделей популяционной биологии, отсылая читателя к соответствующим монографиям, учебниками и учебным пособиям.
Перейдем теперь к построению моделей иного типа.