

Уравнение Ферхюльста (логистическое уравнение)
В качестве примера изложенного подхода рассмотрим так называемое логистическое уравнение или уравнение Ферхюльста, предложенное в 1838 году. Логистическое уравнение описывает единственную гомогенную популяцию особей, соревнующихся за ресурсы, чьё количество ограничено. Современное экспериментальное обоснование этого уравнения относится к области микробиологии, где исходные предположения оказываются наиболее естественными. Исследования такого рода активно проводились в первой половине ХХ века.
В частности, была изучена динамика популяции дрожжей Saccharomyces Cervesia (пивные дрожжи) в растворе сахара. Эксперимент начинается с малого количества дрожжей и в начальной стадии их количество возрастало по экспоненте. Позднее количество сахара уменьшилось и дрожжи начинали конкурировать за ресурсы (сахар). Вторым компонентом конкурентной борьбы может быть отравление дрожжей алкоголем.
Так как рассматриваемая модель хорошо известна и достаточна проста, то мы опишем её очень кратко.
Состояние системы в данный момент времени мы будем описывать концентрацией дрожжей . В системе одновременно протекают два процесса: рождение и смерть. Из-за конкуренции за ресурсы (сахар) скорость рождения уменьшается, а скорость смерти возрастает при увеличении размера популяции. Простейшим предположением являются линейные зависимости:
![]() (4.8)
(4.8)
где
![]() – скорость рождения,
– скорость рождения,
![]() – скорость смерти. Так как
– скорость смерти. Так как
![]() ,
,
то,
введя обозначения
![]() ,
,
![]() ,
получим следующую модель
,
получим следующую модель
![]() .
(4.9)
.
(4.9)
Полученное уравнение называется логистическим уравнением или уравнением Ферхюльста (или логистическим уравнением с непрерывным временем). Конечно, это крайне упрощённая модель, однако, в определённой области параметров она довольно неплохо аппроксимирует экспериментальные результаты. Логистическое уравнение (4.9) легко решается аналитически:
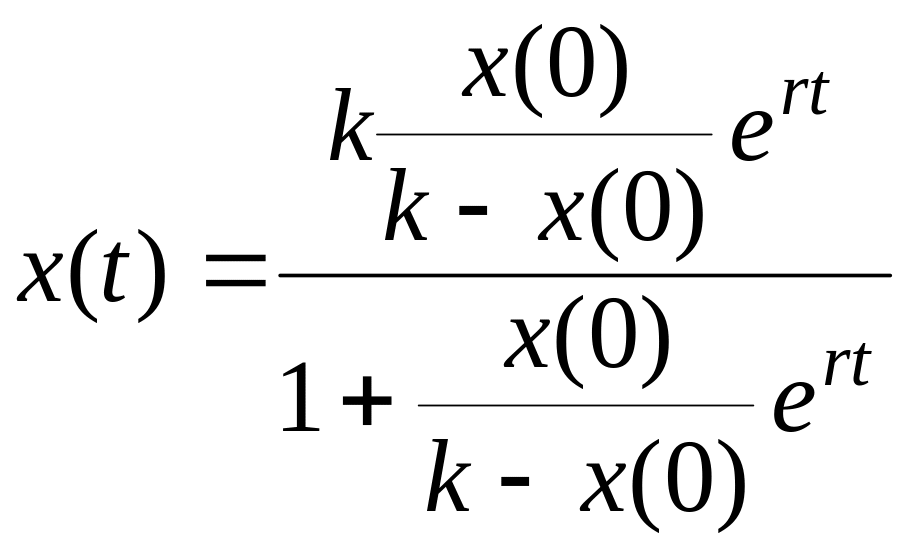 ,
(4.10)
,
(4.10)
Из
уравнения (4.9) легко находятся стационарные
точки
![]() и
и
![]() .
Мы будем рассматривать случай
.
Мы будем рассматривать случай
![]() ,
что соответствует модели, в которой
скорость рождения больше скорости
смерти в отсутствии конкуренции (избыток
ресурсов). Из
следует, что и
,
что соответствует модели, в которой
скорость рождения больше скорости
смерти в отсутствии конкуренции (избыток
ресурсов). Из
следует, что и
![]() .
Рассмотрим устойчивость этих стационарных
точек. Для этого в соответствии с
изложенной теорией рассмотрим знак
производной
.
Рассмотрим устойчивость этих стационарных
точек. Для этого в соответствии с
изложенной теорией рассмотрим знак
производной
![]()
![]() .
(4.11)
.
(4.11)
При
![]() имеем
имеем
![]() (так как
)
и стационарная точка
неустойчива. При
(так как
)
и стационарная точка
неустойчива. При
![]() имеем
имеем
![]() и так как
стационарная точка
устойчива. Качественное поведение
кинетических кривых может быть описано
следующим образом: если
и так как
стационарная точка
устойчива. Качественное поведение
кинетических кривых может быть описано
следующим образом: если
![]() ,
то
монотонно убывающая функция, асимптотически
стремящаяся к
;
если
,
то
монотонно убывающая функция, асимптотически
стремящаяся к
;
если
![]() ,
то
монотонно возрастающая функция без
точек перегиба, асимптотически стремящаяся
к
;
если
,
то
монотонно возрастающая функция без
точек перегиба, асимптотически стремящаяся
к
;
если
![]() ,
то
монотонно возрастающая функция с точкой
перегиба. Соответствующие кинетические
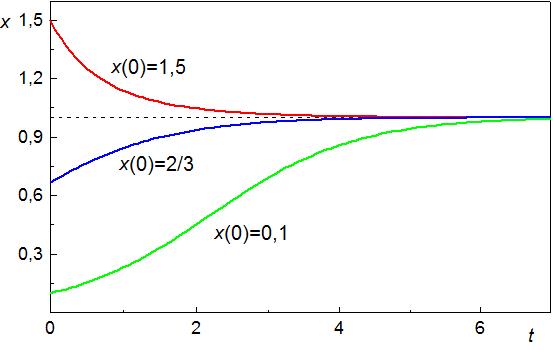
кривые показаны на рис. 4.3.
,
то
монотонно возрастающая функция с точкой
перегиба. Соответствующие кинетические
кривые показаны на рис. 4.3.
Популяционная модель с дискретным временем
Теперь рассмотрим аналогичную модель популяции, но с дискретным временем. Дискретность времени в динамике популяций возникает достаточно естественно: сезонность размножения, световой цикл в течении суток и т.д.
Достаточно общий вид уравнения с дискретным временем может быть записан как
![]() ,
(4.12)
,
(4.12)
где
![]() – численность популяции в
-й
момент времени и т.д.;
– некоторая функция.
– численность популяции в
-й
момент времени и т.д.;
– некоторая функция.
|
Рис. 4.3. Кинетические кривые уравнения Ферхюльста при различных начальных условиях х(0). Время t и объем популяции х в условных единицах. |
Простейший вид уравнение (4.12) принимает в случае, если численность каждого следующего поколения в популяции зависит только от численности предыдущего поколения. Это справедливо для многих видов насекомых, а также некоторых видов рыб, птиц и зоопланктона. Можно сказать, что в этом случае поколения не перекрываются во времени. Уравнение (4.12) принимает следующий вид:
![]() .
(4.13)
.
(4.13)
Удивительно, что даже это простейшее уравнение демонстрирует исключительно разнообразное поведение: монотонность; затухающие колебания; незатухающие колебания с различным периодом; динамический хаос.
В
качестве примера приведём дискретное
логистическое уравнение, отталкиваясь
от непрерывного уравнения (4.9). Заменив
![]() на
на
![]() ,
где
,
где
![]() ,
,
![]() ,
получим дискретный аналог уравнения
(4.9):
,
получим дискретный аналог уравнения
(4.9):
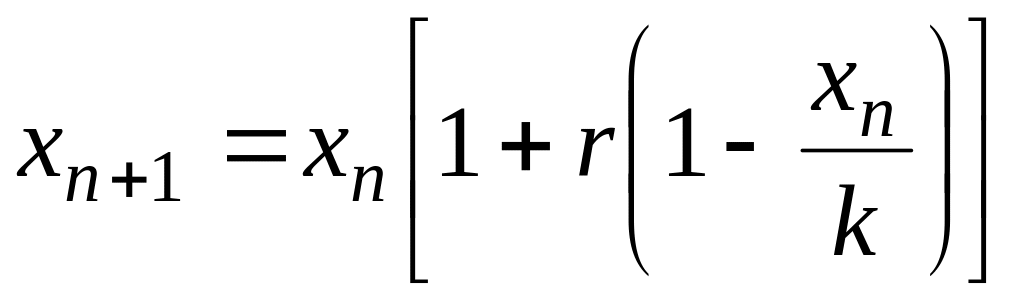 ,
(4.14)
,
(4.14)
Полученное
уравнение, корректное с математической
точки зрения, некорректно с биологической
точки зрения, так как, если
![]() ,
то из уравнения (4.14) следует, что
,
то из уравнения (4.14) следует, что
![]() .
У непрерывного уравнения Ферхюльста
этот недостаток отсутствует. Чтобы от
него избавиться, необходимо модифицировать
правую часть уравнения (4.14). Одна из
возможных модификаций была предложена
для численности популяции насекомых и
рыб:
.
У непрерывного уравнения Ферхюльста
этот недостаток отсутствует. Чтобы от
него избавиться, необходимо модифицировать
правую часть уравнения (4.14). Одна из
возможных модификаций была предложена
для численности популяции насекомых и
рыб:
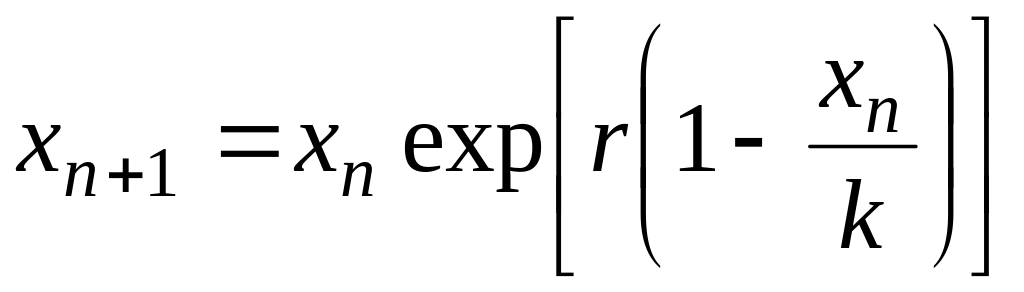 .
(4.15)
.
(4.15)
Для
этого уравнения (4.15) существует устойчивое
стационарное состояние при
![]() .
Значения
.
Значения
![]() стремятся к нему монотонно при
стремятся к нему монотонно при
![]() .
При
.
При
![]() мы видим затухающее колебания. При
мы видим затухающее колебания. При
![]() возникают двухточечные циклы. При
возникают двухточечные циклы. При
![]() последовательно появляются циклы
периода
последовательно появляются циклы
периода
![]() При
При
![]() появляется динамический хаос. Заметим,
что численные значения
появляется динамический хаос. Заметим,
что численные значения
![]() и
и
![]() являются
приближёнными.
являются
приближёнными.
Аналогичными свойствами обладает и исходная модель (4.14) при несколько других значениях концов интервалов. Подобное поведение называется каскадом бифуркаций удвоения периода.
В качестве примера
на рис. 4.4 приведена бифуркационная
диаграмма классического логистического
отображения. По оси абсцисс отложено
значение параметра
,
а по оси ординат – значение
при
![]() .
При
.
При
![]() значения отображения расходятся при
любых начальных значениях.
значения отображения расходятся при
любых начальных значениях.
|
Рис. 4.4. Бифуркационная диаграмма классического логистического отображения
|
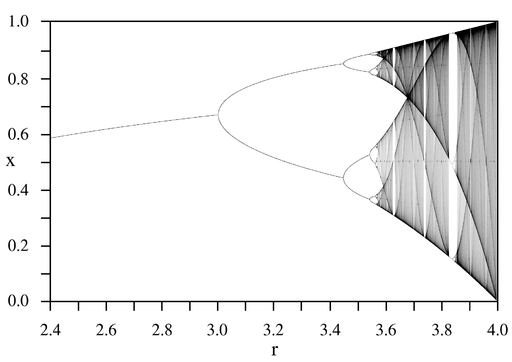
В настоящее время вопрос о том, наблюдается ли обсуждаемое хаотическое поведение в реальных экологических системах, остаётся открытым. Основная проблема состоит в том, что в подавляющем большинстве экологических систем существуют сильные стохастические возмущения (погода, температура и т.д.). По этой причине практически невозможно понять, является ли хаотичность данных следствием нерегулярных возмущений или следствием хаотической динамики. В искусственно созданных популяциях насекомых, в которых очень жестко поддерживались постоянные условия, удалось показать, что динамический хаос действительно имеет место в экологических системах и не только качественно, но и количественно описывается соответствующими уравнениями.
Рассмотрим теперь более сложные экологические системы, для описания которых требуется более чем одна переменная. Общая теория таких систем хорошо разработана для случая двух переменных, к рассмотрению которого мы и переходим.
Вообще, динамика популяций, которую мы начали рассматривать, является классическим объектом биоматематики и математического моделирования в биологии. Идеи, высказанные Лоткой и Вольтеррой в первой половине ХХ века и знаменитый пример с кроликами и лисами, являются основой для изучения взаимодействия различных биологических видов или отдельных субъектов. Отметим одно принципиальное отличие биологических систем от физических. В физических системах значительную роль играют всевозможные симметрии и законы сохранения (массы, энергии, импульсов и т.д.), что существенно облегчает описание протекающих процессов. В биологии, к сожалению, это не так и мощные методы, разработанные в физике, не могут быть использованы при исследовании биологических систем без их существенного модифицирования.