

17. Темпы роста популяций. Факторы, влияющие на численности популяци: зависящие и не зависящие от плотности.
Графики роста числа членов любой природной популяции в новых для нее условиях сильно отличаются от экспоненты. Кривая после подъема разной степени крутизны поворачивает параллельно горизонтальной оси, чем знаменует установление некоторой предельной численности популяции, которая затем поддерживается в течение более или менее длительного времени (рис. 1). Такой ход кривой показывает, что в природе какие
то причины сдерживают чрезмерный рост популяции, не давая ей реализовать свой биотический потенциал, и ограничивают ее численность определенными пределами.
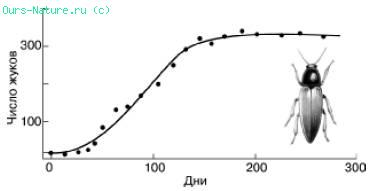
Рис. 1. Численность жуков в культуре, начатой с одной пары Rhizopertha dominica, в 10 г пшеницы (по Дж. Варли, 1978)Пшеницу каждую неделю просеивали и дополняли до 10 гИзменения темпов роста популяции могут быть различными. В одном случае темп роста с самого начала высок и постоянен независимо от нарастающей плотности, что соответствует лавинообразному, по экспоненте, увеличению численности популяции. При достижении же определенной плотности населения темп роста падает сразу до нуля. Это означает, что популяция резко прекращает воспроизводство.
В природе такой тип захвата среды встречается у видов, для которых жизненно важна скорость овладения ресурсами, например у нематод, клещей, некоторых насекомых, обитающих в быстро меняющих качество гниющих растительных остатках, кучках навоза копытных и т. п. Мелкие нематоды, населяющие конский и коровий навоз, могут осуществить свой жизненный цикл за несколько часов, тогда как близкие им виды, но обитающие в других, более постоянных условиях, развиваются от двух до трех недель. Скорость овладения средой имеет значение и для предупреждения конкуренции со стороны других претендентов. После массового размножения и быстрого потребления доступных ресурсов в популяциях подобных видов размножение прекращается и особи расселяются путем перелета на насекомых.
В другом случае изменения темпов роста популяций находятся в обратной зависимости от плотности. Разреженная популяция в благоприятных условиях быстро наращивает свою численность, но чем больше становится в ней особей, тем меньше доля очередного приплода, пока скорость воспроизводства не сравняется со смертностью (рис. 2). Тогда темп роста популяции снижается до нуля, а общая численность стабилизируется в соответствии с доступными для популяции ресурсами. Такой характер становления новых популяций свойствен в основном видам, у которых успех в размножении при низком уровне численности не лимитируется обязательностью группового образа жизни, необходимостью встречи полов и другими причинами (например, у мелких партеногенетических ракообразных, растений с апомиксисом и т. д.). Однако у большинства видов наиболее высокий темп роста популяций отмечается лишь при определенной оптимальной плотности. Если популяция сильно разрежена, это затрудняет встречу полов, защиту молодняка, проявление эффекта группы у животных, опыление у растений, поэтому население растет вначале очень медленно.
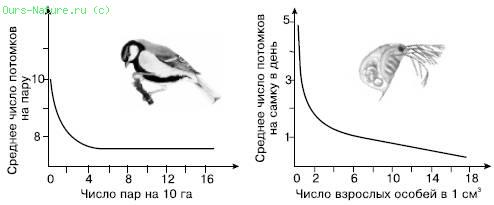
Рис. 2. Зависимость рождаемости от плотности в лабораторной популяции дафний и в дикой популяции большой синицы (по Ю. Одуму, 1975)
Как известно, появление потомства прежде всего зависит от числа производителей – особей, находящихся в генеративном возрастном состоянии. Вначале рост числа производителей сопровождается медленным ростом численности популяции, затем, в некотором интервале плотностей, зависимость выражена чрезвычайно резко, и даже небольшое увеличение доли производителей вызывает стремительный рост популяции, пока она не достигнет определенного уровня, который в последующем уже не меняется, сколько бы ни увеличивалась репродуктивная часть населения.
Такая зависимость была впервые предсказана французским математиком Верхюльстом в середине XIX века для населения людей, а позднее доказана англичанином Перлом (1925) для популяций животных в среде, где пищевые ресурсы имеют определенный предел восполнимости.
Установление определенного уровня плотности популяций после некоторого периода роста вовсе не означает, что в популяциях больше не происходит количественных изменений. Напротив, любое население всегда динамично и постоянно подвержено колебаниям численности, однако размах всех суточных, сезонных и годовых изменений популяций все же намного меньше теоретически возможного, соответствующего реализации всего биотического потенциала. Колебания численности совершаются с разным размахом вокруг некоторой средней величины, которая и соответствует горизонтальной части кривой на графике роста и стабилизации численности популяции.
Высокий потенциал размножения играет большую роль в выживании видов. Популяции, сведенные к низкому уровню численности, могут быстро восстановиться при благоприятной перемене условий. Некоторые виды только массовым размножением могут противостоять выеданию их различными потребителями или угрозе вытеснения конкурентами. Высокая размножаемость способствует быстрому освоению видом новых пространств.
Однако безграничное размножение таит в себе и большую опасность для любой популяции, так как может привести к быстрому подрыву ресурсов среды, нехватке пищи, убежищ, пространства и т. п., что неминуемо повлечет за собой общее ослабление популяции. Перенаселенность настолько неблагоприятна для любого вида, что в ходе эволюции у разных форм выработались в результате естественного отбора самые разнообразные механизмы, способствующие предотвращению избытка особей и поддержанию определенного уровня плотности популяций.