

Скорость реакция цикла Кребса определяется энергетическими потребностями клетки.
Скорость реакций цикла Кребса коррелирует с интенсивностью процесса тканевого дыхания и связанного с ним окислительного фосфорилирования. В свою очередь, функцио(дыхательного контроля): чем меньше это соотношение, тем интенсивнее идёт тканевое дыхание и эффективнее генерируется электро-химический потенциал, т. е. увеличивается выработка АТФ. Ускорение тканевого дыхания приводит к более быстрому потреблению субстратов цикла Кребса, которые одновременно являются также субстратами тканевого дыхания.
Непосредственно регулируется активность дегидрогеназ цикла Кребса, которая зависит от величины отношения НАДН∙Н+/ НАД+: при недостатке НАД+ их активность снижается, поскольку именно окисленная форма кофермента является акцептором водородов для субстратов лимоннокислого цикла. Помимо этого, существует аллостерический механизм регуляции: для изоцитрадегидрогеназы аллостерическими ингибиторами являются АТФ и НАДН∙Н+, активатором – АДФ. Сукцинатдегидрогеназа угнетается избытком субстрата.
1. Пентозофосфатный путь окисления глюкозы обслуживает восстановительные синтезы в клетке.
Как видно из названия, в этом пути образуются столь необходимые клетке пентозофосфаты. Поскольку образование пентоз сопровождается окислением и отщеплением первого углеродного атома глюкозы, то этот путь называется также апотомическим (apex – вершина).
Пентозофосфатный путь можно разделить 2 части: окислительную и неокислительную. В окислительной части, включающей 3 реакции, образуются НАДФН∙Н+ и рибулозо-5-фосфат. В неокислительной части рибулозо-5-фосфат превращается в различные моносахариды с 3, 4, 5, 6, 7 и 8-ью атомами углерода; конечными продуктами являются фруктозо-6-фосфат и 3-ФГА.
Окислительная часть.
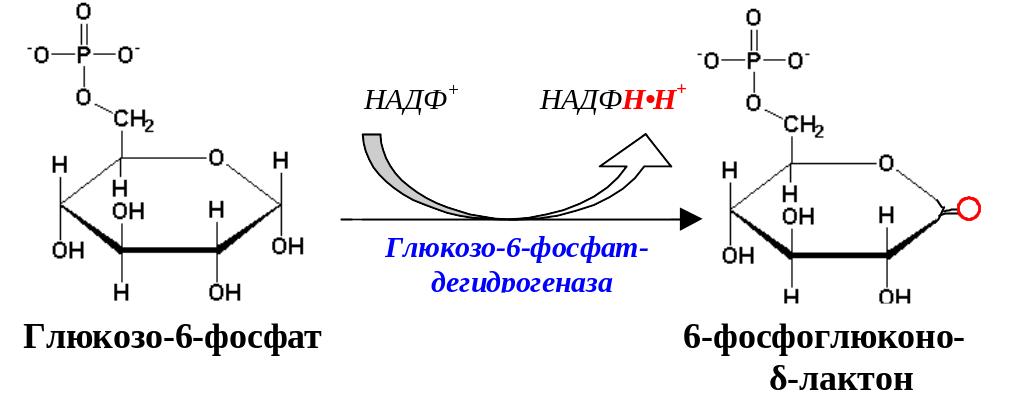
Первая реакция – дегидрирование глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой с образованием δ-лактона 6-фосфоглюконовой кислоты и НАДФН∙Н+ (НАДФ+ – кофермент глюкозо-6-фосфатдегидрогеназы).
Вторая реакция – гидролиз 6-фосфоглюконолактона глюконолактонгидролазой. Продукт реакции – 6-фосфоглюконат.
глюконолактонгидролаза
 6-фосфоглюконолактон
6-фосфоглюконат
6-фосфоглюконолактон
6-фосфоглюконат
Н2О
Третья реакция – дегидрирование и декарбоксилирование 6-фосфоглюко-нолактона ферментом 6-фосфоглюконатдегидрогеназой, коферментом которого является НАДФ+ . В ходе реакции восстанавливается кофермент и отщепляется 1С глюкозы с образованием рибулозо-5-фосфата.
СООН
│
 Н–С–ОН
СН2ОН
Н–С–ОН
СН2ОН
│ СО2 │
Н–С–ОН С=О
│ │
Н–С–ОН 6-фосфоглюконатдегидрогеназа Н–С–ОН
│ (декарбоксилирующая) │
Н–С–ОН Н–С–ОН
│ │
СН2ОРО3Н2 СН2ОРО3Н2
6-фосфоглюконат Рибулозо-5-фосфат
Неокислительная часть.
В отличие от первой, окислительной, все реакции этой части пентозофосфатного пути обратимы.
Рибулозо-5-фосфат может изомеризоваться (фермент – кетоизомераза) в рибозу-5-фосфат и эпимеризоваться (фермент – епимераза) в ксилулозо-5-фосфат. Далее следуют 2 типа реакций: транскетолазная и трансальдолазная.
Транскетолаза (кофермент – тиаминпирофосфат) отщепляет 2С-фрагмент и переносит его на другие сахара (см. схему). Трансальдолаза способна переносить 3С-фрагменты.
В реакцию вначале вступают рибозо-5-фосфат и ксилулозо-5-фосфат. Это – транскетолазная реакция: переносится 2С-фрагмент от ксилулозо-5-фос-фата на рибозо-5-фосфат.
Значение пентозофосфатного пути окисления глюкозы
Ферменты пентозофосфатного пути локализуются в цитоплазме. В тканях, синтезирующих стероиды или жирные кислоты (для чего необходим НАДФН∙Н+), окислительная часть пути протекает весьма интенсивно. К таким тканям относятся: надпочечники, печень, жировая ткань, лактирующая молочная железа. Эритроциты нуждаются в коферменте НАДФН∙Н+ для восстановления глутатиона (трипептид). Совместно с витамином С восстановленный глутатион играет основную роль в предупреждении образования метгемоглобина: глутатион-SH является активной частью глутатионпероксидазы, устраняющей токсическое влияние пероксида водорода и других перекисей, окисляющих железо гемоглобина и нарушающих его кислородтранспортную функцию.

глутатион –S-S-глутатион
НАДФН∙ Н+ Н2О
Глутатионредуктаза Глутатионпероксидаза
НАДФ+ Н2О2