

1 88 Глава 8
Рис. 126. В типичном рецептивном поле типа 1 центр посылает возбуждающие сигналы от красных колбочек, а периферия — тормозящие сигналы от зеленых колбочек.
Следующий шаг состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки; это мы и сделали в 1966 году совместно с Торстеном Визелом. У рецептивных полей большинства оппонентно-цветовых клеток де Валуа обнаружилась удивительная организация, которая до сих пор ставит нас в тупик. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию; центр мог быть типа on или off. В типичном случае центр поля представлен исключительно красными, а тормозящая периферия — исключительно зелеными колбочками. Поэтому при красном свете как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порождают сильное торможение с off-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа.
Хотя нашим первым впечатлением было, что такая клетка из центра % должна получать входные сигналы от красных, а с периферии — от зеленых колбочек, теперь кажется вероятным, что полное рецептивное поле включает две перекрывающиеся системы, как это показано на рис. 127. Как красные, так и зеленые колбочки распределены по довольно широкой круговой области, причем их количество максимально в центре и убывает по мере удаления от него. В центре красные колбочки сильно преобладают, но к периферии число их падает значительно быстрее числа зеленых колбочек. Поэтому небольшое пятнышко, вспыхивающее в центре и содержащее длинноволновую компоненту, будет весьма мощным стимулом для красной системы; даже если оно стимулирует и зеленые колбочки, их число по сравнению с общим числом связанных с клеткой зеленых колбочек будет слишком мало, чтобы составить какую-либо конкуренцию красной системе. Те же соображения приложимы и к клеткам с центром и периферией, описанным в главе 3, рецептивные поля которых тоже должны состоять из двух оппонентных круговых перекрывающихся зон с разными формами кривых зависимости чувствительности от координаты. Таким образом, периферия, вероятно, имеет не кольцеобразную, как думали вначале, а круговую форму. В отношении этих оппонентно-цветовых клеток обезьян предполагается (правда, без достаточных данных), что периферические области отражают вклад горизонтальных клеток.
Реакция на диффузный свет — в данном случае on-реакции на красный, off-

ЦВЕТОВОЕ ЗРЕНИЕ
189
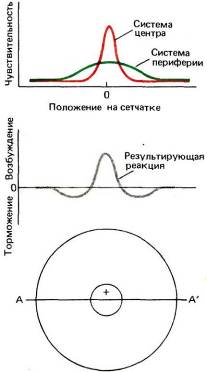
Рис. 127. Графики вверху отражают зависимость чувствительности нейрона (измеренной, например, по реакции на неподвижное очень маленькое световое пятнышко) от положения стимула на сетчатке вдоль линии АА, проходящей через центр рецептивного поля. Для клетки с r-центром и g~-nepn(|)epHefi красное пятнышко дает островершинную кривую, а зеленое — значительно более широкую кривую. Нижний график отображает реакцию на белый или желтый свет, стимулирующий обе оппонентные системы, так что происходит вычитание вкладов двух систем. При этом красные колбочки доминируют в центре, что приводит здесь к on-реакциям, а зеленые — на периферии, что приводит к off-реакциям.
реакции на синий и зеленый и отсутствие реакции на белый свет — ясно показывают, что такая клетка должна регистрировать информацию о цвете. Но реакции на определенного рода белые края и отсутствие реакций на диффузный свет убеждают в том, что эта клетка связана также и с восприятием формы. Мы называем эти цветооппонентные клетки с центром и периферией клетками «типа 1».
Наружное коленчатое тело обезьяны, если вспомнить сведения из главы 4, содержит шесть слоев, причем четыре верхних слоя содержат множество мелких клеток, а два нижних — меньшее число крупных клеток. Мы находим клетки описанного выше типа 1 в верхних, или мелкоклеточных, слоях. Они различаются по типу колбочек, входящих в их центральную и периферийную системы, и по характеру центра, который может быть возбуждающим или тормозящим. Пример, представленный на рис. 126, мы можем обозначить как r+g~ (с красным центром и зеленой периферией). Среди подтипов клеток, получающих входы от красных и зеленых колбочек, мы находим все четыре возможных варианта: r+ g~, r~g+, g+r", g~r+. Вторая группа клеток получает входы от синих колбочек из центра поля и от комбинации красных и зеленых (или, быть может, только от зеленых) — с периферии. Мы называем такие