

Глава 7
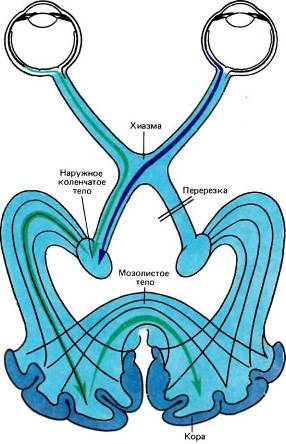
Рис. 100. Уиттеридж в своем эксперименте произвел перерезку правого зрительного тракта. Теперь для того, чтобы информация поступала в зрительную кору правого полушария, она должна сначала попасть в зрительную кору левого полушария, а затем пройти в другое полушарие через мозолистое тело. Охлаждение какого-либо участка этого пути блокирует передачу нервных импульсов.
полушариями через мозолистое тело должно приводить в итоге к тому, что каждое полушарие будет получать сигналы от области несколько большей, чем половина поля зрения. Иными словами, за счет связей через мозолистое тело произойдет перекрывание полуполей, проецируемых в два полушария. Именно это мы и обнаружили. С помощью двух электродов, введенных в область коры у границы полей 17 и 18 в каждом из полушарий, нам нередко удавалось регистрировать активность клеток, рецептивные поля которых взаимно перекрывались на несколько угловых градусов.
Т. Визел и я вскоре произвели микроэлектродные отведения непосредственно от той зоны мозолистого тела (в самой задней его части), где имеются волокна, связанные со зрительной системой. Мы нашли, что почти все волокна, которые мы могли активировать зрительными стимулами, отвечали в точности так же, как и обычные нейроны поля 17, т. е. проявляли свойства как простых, так и сложных клеток, избирательно чувствительных к ориентации стимула и обычно отвечавших на стимуляцию обоих глаз. Во всех этих случаях рецептивные поля располагались очень близко к средней вертикали ниже или выше (либо на уровне) точки фиксации, как показано на рис. 101.
Пожалуй, наиболее изящной нейрофизиологической демонстрацией роли
МОЗОЛИСТОЕ ТЕЛО И СТЕРЕОСКОПИЧЕСКОЕ ЗРЕНИЕ
149
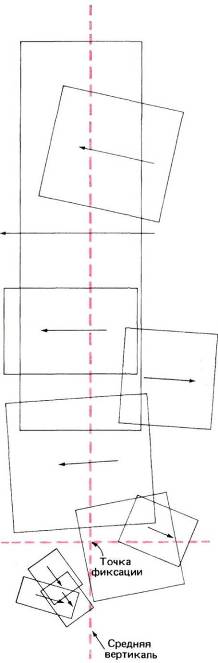
Рис. 101. Рецептивные поля волокон мозолистого тела лежат очень близко к средней вертикали поля зрения. Все эти поля были выявлены путем регистрации активности десяти волокон у одной кошки.
мозолистого тела стала работа Дж. Берлукки и Дж. Риццолатти из г. Пизы, выполненная в 1968 году. Перерезав зрительную хиазму по средней линии, они регистрировали ответы в поле 17 вблизи границы с полем 18, отыскивая те клетки, которые могли активироваться бинокулярно. Ясно, что любая бинокулярная клетка этой области в правом полушарии должна получать входные сигналы как прямо от правого глаза (через НКТ), так и от левого глаза и левого полушария через мозолистое тело. Как выяснилось, рецептивное поле каждой
150 Глава 7
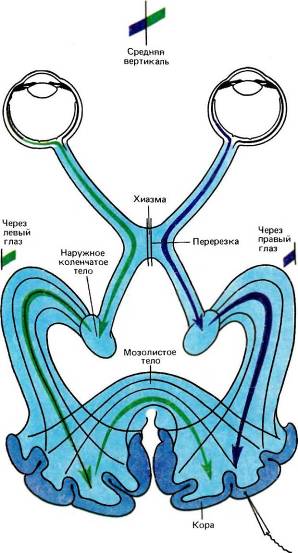
Рис. 102. Берлукки и Риццолатти в своем эксперименте изящно продемонстрировали не только функцию зрительной части волокон мозолистого тела, но и высокую специфичность межполушарных связей между клетками, сходными по ориентации и расположению рецептивных полей. Берлукки и Риццолатти перерезали у кошки хиазму по средней линии, так что левый глаз стал передавать информацию только в левое полушарие, причем эта информация касалась только правой половины поля зрения. Точно так же правый глаз стал снабжать информацией только правое полушарие, и эта информация относилась только к левой половине поля зрения. После перерезки регистрировались ответы тех клеток, рецептивные поля которых в нормальных условиях пересекали бы среднюю вертикаль. Оказалось, что рецептивные поля таких клеток расщепляются по этой вертикали; правая часть связана теперь с левым глазом, а левая — с правым глазом.
МОЗОЛИСТОЕ ТЕЛО И СТЕРЕОСКОПИЧЕСКОЕ ЗРЕНИЕ 151
бинокулярной клетки захватывало среднюю вертикаль сетчатки, причем та его часть, которая относится к левой половине поля зрения, доставляла информацию от правого глаза, а та, которая заходит в правую половину, — от левого глаза. Другие свойства клеток, исследованные в этом эксперименте, включая ориентационную избирательность, оказались идентичными (рис. 102).
Полученные результаты ясно показали, что мозолистое тело связывает клетки друг с другом таким образом, чтобы их рецептивные поля могли заходить и вправо, и влево от средней вертикали. Таким образом, оно как бы склеивает две половины изображения окружающего мира. Чтобы лучше себе это представить, предположим, что изначально кора нашего мозга образовалась как одно целое, не разделенное на два полушария. В этом случае поле 17 имело бы вид одного непрерывного слоя, на который отображалось бы все зрительное поле. Тогда соседние клетки для реализации таких свойств как, например, чувствительность к движению и ориентационная избирательность, должны были бы иметь, разумеется, сложную систему взаимных связей. Теперь вообразим, что «конструктор» (будь то бог, или, скажем, естественный отбор) решил, что так дальше оставлять нельзя — отныне половина всех клеток должна образовать одно полушарие, а другая половина — другое полушарие. Что тогда нужно сделать со всем множеством межклеточных связей, если две совокупности клеток должны теперь отодвинуться друг от друга? По-видимому, можно просто растянуть эти связи, образовав из них часть мозолистого тела. Для того чтобы устранить задержку при передаче сигналов по такому длинному пути (у человека примерно 12—15 сантиметров), нужно увеличить скорость передачи, снабдив волокна мйелйновой оболочкой. Разумеется, на самом деле ничего такого в процессе эволюции не происходило; задолго до того, как возникла кора, мозг уже имел два отдельных полушария.
Эксперимент Берлукки и Риццолатти, на мой взгляд, дал одно из наиболее ярких подтверждений удивительной специфичности нейронных связей. Клетка, показанная на рис. 108 (около кончика электрода) и, вероятно, миллион других подобных клеток, соединенных через мозолистое тело, приобретают свою ориентационную избирательность как за счет локальных связей с соседними клетками, так и за счет связей, идущих через мозолистое тело из другого полушария от клеток с такой же ориентационной чувствительностью и сходным расположением рецептивных полей (сказанное относится и к другим свойствам клеток, таким как дирекциональная специфичность, способность реагировать на концы линий, а также сложность). Каждая из клеток зрительной коры, имеющих связи через мозолистое тело, должна получать входные сигналы от клеток другого полушария с точно такими же свойствами. Мы знаем множество фактов, указывающих на избирательность соединений в нервной системе, но я думаю, что данный пример — наиболее яркий и убедительный.
Рассмотренные выше аксоны клеток зрительной коры составляют лишь небольшую долю всех волокон мозолистого тела. На соматосенсорной коре проводились эксперименты с использованием аксонного транспорта, аналогичные описанным в предыдущих главах опытам с инъекцией радиоактивной аминокислоты в глаз. Их результаты показывают, что мозолистое тело таким же образом связывает те участки коры, которые активируются кожными и суставными рецепторами, расположенными вблизи средней линии тела на туловище и голове, но не связывает корковые проекции конечностей.
Каждая область коры соединяется с несколькими или даже многими другими областями коры того же полушария. Например, первичная зрительная кора связана с полем 18 (зрительной зоной 2), с медиальной височной областью (зоной МТ), со зрительной зоной 4 и еще с одной или двумя областями. Многие участки коры имеют также связи с несколькими областями другого полушария,